Thank you for visiting nature.com. You are using a browser version with limited support for CSS. To obtain the best experience, we recommend you use a more up to date browser (or turn off compatibility mode in Internet Explorer). In the meantime, to ensure continued support, we are displaying the site without styles and JavaScript.
- View all journals
- Explore content
- About the journal
- Publish with us
- Sign up for alerts
- Review Article
- Published: 24 September 2021

Abiotic stress responses in plants
- Huiming Zhang ORCID: orcid.org/0000-0003-0695-3593 1 ,
- Jianhua Zhu 2 ,
- Zhizhong Gong 3 , 4 &
- Jian-Kang Zhu ORCID: orcid.org/0000-0001-5134-731X 1
Nature Reviews Genetics volume 23 , pages 104–119 ( 2022 ) Cite this article
49k Accesses
1018 Citations
87 Altmetric
Metrics details
- Agricultural genetics
- Molecular biology
- Plant genetics
- Plant sciences
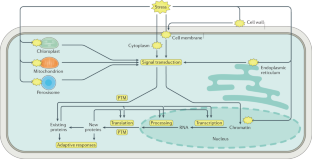
Plants cannot move, so they must endure abiotic stresses such as drought, salinity and extreme temperatures. These stressors greatly limit the distribution of plants, alter their growth and development, and reduce crop productivity. Recent progress in our understanding of the molecular mechanisms underlying the responses of plants to abiotic stresses emphasizes their multilevel nature; multiple processes are involved, including sensing, signalling, transcription, transcript processing, translation and post-translational protein modifications. This improved knowledge can be used to boost crop productivity and agricultural sustainability through genetic, chemical and microbial approaches.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout
Similar content being viewed by others

Plant hormone regulation of abiotic stress responses

Metabolic, physiological and anatomical responses of soybean plants under water deficit and high temperature condition

Cross-stress gene expression atlas of Marchantia polymorpha reveals the hierarchy and regulatory principles of abiotic stress responses
Bailey-Serres, J., Parker, J. E., Ainsworth, E. A., Oldroyd, G. E. D. & Schroeder, J. I. Genetic strategies for improving crop yields. Nature 575 , 109–118 (2019).
CAS PubMed PubMed Central Google Scholar
Zhang, H., Zhao, Y. & Zhu, J. K. Thriving under stress: how plants balance growth and the stress response. Dev. Cell 55 , 529–543 (2020).
CAS PubMed Google Scholar
Zhu, J. K. Abiotic stress signaling and responses in plants. Cell 167 , 313–324 (2016). This Review proposes the concept of dispersed stress sensing in various cell parts .
Chen, X. X. et al. Protein kinases in plant responses to drought, salt, and cold stress. J. Integr. Plant Biol. 63 , 53–78 (2021).
Gupta, A., Rico-Medina, A. & Cano-Delgado, A. I. The physiology of plant responses to drought. Science 368 , 266–269 (2020).
Takahashi, F., Kuromori, T., Urano, K., Yamaguchi-Shinozaki, K. & Shinozaki, K. Drought stress responses and resistance in plants: from cellular responses to long-distance intercellular communication. Front. Plant Sci. 11 , 556972 (2020).
PubMed PubMed Central Google Scholar
Yang, Y. & Guo, Y. Unraveling salt stress signaling in plants. J. Integr. Plant Biol. 60 , 796–804 (2018).
Zhang, J. Y., Li, X. M., Lin, H. X. & Chong, K. Crop improvement through temperature resilience. Annu. Rev. Plant Biol. 70 , 753–780 (2019).
Yuan, F. et al. OSCA1 mediates osmotic-stress-evoked Ca 2+ increases vital for osmosensing in Arabidopsis . Nature 514 , 367–371 (2014).
Jojoa-Cruz, S. et al. Cryo-EM structure of the mechanically activated ion channel OSCA1.2. eLife 7 , e41845 (2018).
Liu, X., Wang, J. & Sun, L. Structure of the hyperosmolality-gated calcium-permeable channel OSCA1.2. Nat. Commun. 9 , 5060 (2018).
Maity, K. et al. Cryo-EM structure of OSCA1.2 from Oryza sativa elucidates the mechanical basis of potential membrane hyperosmolality gating. Proc. Natl Acad. Sci. USA 116 , 14309–14318 (2019).
Hamilton, E. S. et al. Mechanosensitive channel MSL8 regulates osmotic forces during pollen hydration and germination. Science 350 , 438–441 (2015).
Hamilton, E. S. & Haswell, E. S. The tension-sensitive ion transport activity of MSL8 is critical for its function in pollen hydration and germination. Plant Cell Physiol. 58 , 1222–1237 (2017).
Jiang, Z. et al. Plant cell-surface GIPC sphingolipids sense salt to trigger Ca 2+ influx. Nature 572 , 341–346 (2019). This work demonstrates an important role for MOCA1-dependent GIPC production in plant response to high salinity .
Rennie, E. A. et al. Identification of a sphingolipid α-glucuronosyltransferase that is essential for pollen function in Arabidopsis . Plant Cell 26 , 3314–3325 (2014).
Laohavisit, A. et al. Salinity-induced calcium signaling and root adaptation in arabidopsis require the calcium regulatory protein annexin1. Plant Physiol. 163 , 253–262 (2013).
Ma, L. et al. The SOS2–SCaBP8 complex generates and fine-tunes an AtANN4-dependent calcium signature under salt stress. Dev. Cell 48 , 697–709.e5 (2019).
Feng, W. et al. The FERONIA receptor kinase maintains cell-wall integrity during salt stress through Ca 2+ signaling. Curr. Biol. 28 , 666–675.e5 (2018).
Zhao, C. et al. Leucine-rich repeat extensin proteins regulate plant salt tolerance in Arabidopsis . Proc. Natl Acad. Sci. USA 115 , 13123–13128 (2018). Together with Feng et al. (2018), this work supports the critical roles of the LRX–RALF–FER regulatory module in salt sensing and salt tolerance .
Sangwan, V., Orvar, B. L., Beyerly, J., Hirt, H. & Dhindsa, R. S. Opposite changes in membrane fluidity mimic cold and heat stress activation of distinct plant MAP kinase pathways. Plant J. 31 , 629–638 (2002).
Cui, Y. et al. Cyclic nucleotide-gated ion channels 14 and 16 promote tolerance to heat and chilling in rice. Plant Physiol. 183 , 1794–1808 (2020).
Liu, Q. et al. The calcium transporter ANNEXIN1 mediates cold-induced calcium signaling and freezing tolerance in plants. EMBO J. 7 , e104559 (2020). This study reports cold-induced Ca 2+ influx through the Ca 2+ transporter AtANN1 .
Google Scholar
Ma, Y. et al. COLD1 confers chilling tolerance in rice. Cell 160 , 1209–1221 (2015).
Wang, J. et al. Transcriptional activation and phosphorylation of OsCNGC9 confer enhanced chilling tolerance in rice. Mol. Plant 14 , 315–329 (2020).
PubMed Google Scholar
Jiang, B. et al. Cold-Induced CBF–pif3 interaction enhances freezing tolerance by stabilizing the phyB thermosensor in Arabidopsis . Mol. Plant 13 , 894–906 (2020).
Jung, J. H. et al. Phytochromes function as thermosensors in Arabidopsis . Science 354 , 886–889 (2016).
Legris, M. et al. Phytochrome B integrates light and temperature signals in Arabidopsis . Science 354 , 897–900 (2016).
Casal, J. J. & Balasubramanian, S. Thermomorphogenesis. Annu. Rev. Plant Biol. 70 , 321–346 (2019).
Jung, J. H. et al. A prion-like domain in ELF3 functions as a thermosensor in Arabidopsis . Nature 585 , 256–260 (2020). This study identifies a thermosensor protein that displays a direct biophysical (that is, phase transition) response to increasing temperature .
Scharf, K. D., Berberich, T., Ebersberger, I. & Nover, L. The plant heat stress transcription factor (Hsf) family: structure, function and evolution. Biochim. Biophys. Acta 1819 , 104–119 (2012).
Kumar, S. V. & Wigge, P. A. H2A.Z-containing nucleosomes mediate the thermosensory response in Arabidopsis . Cell 140 , 136–147 (2010).
McAinsh, M. R. & Pittman, J. K. Shaping the calcium signature. N. Phytol. 181 , 275–294 (2009).
CAS Google Scholar
Marti, M. C., Stancombe, M. A. & Webb, A. A. R. Cell- and stimulus type-specific intracellular free Ca 2+ signals in arabidopsis. Plant Physiol. 163 , 625–634 (2013).
Hrabak, E. M. et al. The Arabidopsis CDPK–SnRK superfamily of protein kinases. Plant Physiol. 132 , 666–680 (2003).
Tang, R. J., Wang, C., Li, K. & Luan, S. The CBL–CIPK calcium signaling network: unified paradigm from 20 years of discoveries. Trends Plant Sci. 25 , 604–617 (2020).
Tang, R. J. et al. Tonoplast CBL–CIPK calcium signaling network regulates magnesium homeostasis in Arabidopsis . Proc. Natl Acad. Sci. USA 112 , 3134–3139 (2015).
Kim, Y., Park, S., Gilmour, S. J. & Thomashow, M. F. Roles of CAMTA transcription factors and salicylic acid in configuring the low-temperature transcriptome and freezing tolerance of Arabidopsis . Plant J. 75 , 364–376 (2013).
Geiger, D. et al. Activity of guard cell anion channel SLAC1 is controlled by drought-stress signaling kinase–phosphatase pair. Proc. Natl Acad. Sci. USA 106 , 21425–21430 (2009).
Klingler, J. P., Batelli, G. & Zhu, J. K. ABA receptors: the START of a new paradigm in phytohormone signalling. J. Exp. Bot. 61 , 3199–3210 (2010).
Min, M. K. et al. Two clade A phosphatase 2Cs expressed in guard cells physically interact with abscisic acid signaling components to induce stomatal closure in rice. Rice 12 , 37 (2019).
Sirichandra, C. et al. Phosphorylation of the Arabidopsis AtrbohF NADPH oxidase by OST1 protein kinase. FEBS Lett. 583 , 2982–2986 (2009).
Sun, S. J. et al. Protein kinase OsSAPK8 functions as an essential activator of S-type anion channel OsSLAC1, which is nitrate-selective in rice. Planta 243 , 489–500 (2016).
Wang, P. et al. Mapping proteome-wide targets of protein kinases in plant stress responses. Proc. Natl Acad. Sci. USA 117 , 3270–3280 (2020).
Wu, Q. Q. et al. ZmOST1 mediates abscisic acid regulation of guard cell ion channels and drought stress responses. J. Integr. Plant Biol. 61 , 478–491 (2019).
Ma, Y. et al. Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science 324 , 1064–1068 (2009).
Park, S. Y. et al. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science 324 , 1068–1071 (2009).
Chen, K. et al. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 62 , 25–54 (2020).
Ding, Y. et al. OST1 kinase modulates freezing tolerance by enhancing ICE1 stability in arabidopsis. Dev. Cell 32 , 278–289 (2015). This study demonstrates ABA-independent activation of SnRK2.6/OST1 by cold stress .
Katsuta, S. et al. Arabidopsis Raf-like kinases act as positive regulators of subclass III SnRK2 in osmostress signaling. Plant J. 103 , 634–644 (2020).
Lin, Z. et al. A RAF–SnRK2 kinase cascade mediates early osmotic stress signaling in higher plants. Nat. Commun. 11 , 613 (2020).
Soma, F., Takahashi, F., Suzuki, T., Shinozaki, K. & Yamaguchi-Shinozaki, K. Plant Raf-like kinases regulate the mRNA population upstream of ABA-unresponsive SnRK2 kinases under drought stress. Nat. Commun. 11 , 1373 (2020).
Takahashi, Y. et al. MAP3Kinase-dependent SnRK2-kinase activation is required for abscisic acid signal transduction and rapid osmotic stress response. Nat. Commun. 11 , 12 (2020).
Fabregas, N., Yoshida, T. & Fernie, A. R. Role of Raf-like kinases in SnRK2 activation and osmotic stress response in plants. Nat. Commun. 11 , 6184 (2020). Together with Lin et al. (2020), Soma at al. (2020) and Takahashi et al. ( Nature Communications , 2020), this paper demonstrates that hyperosmotic stress rapidly activates RAFs, leading to phosphorylation and activation of ABA-independent SnRK2s .
Lin, Z. et al. Initiation and amplification of SnRK2 activation in abscisic acid signaling. Nat. Commun. 12 , 2456 (2021).
Gobert, A., Isayenkov, S., Voelker, C., Czempinski, K. & Maathuis, F. J. The two-pore channel TPK1 gene encodes the vacuolar K + conductance and plays a role in K + homeostasis. Proc. Natl Acad. Sci. USA 104 , 10726–10731 (2007).
Isner, J. C., Begum, A., Nuehse, T., Hetherington, A. M. & Maathuis, F. J. M. KIN7 kinase regulates the vacuolar TPK1 K + channel during stomatal closure. Curr. Biol. 28 , 466–472 e4 (2018).
Chen, X. X. et al. Arabidopsis U-box E3 ubiquitin ligase PUB11 negatively regulates drought tolerance by degrading the receptor-like protein kinases LRR1 and KIN7. J. Integr. Plant Biol. 63 , 494–509 (2021).
Yang, T. et al. Calcium/calmodulin-regulated receptor-like kinase CRLK1 interacts with MEKK1 in plants. Plant Signal. Behav. 5 , 991–994 (2010).
Zhao, C. et al. MAP kinase cascades regulate the cold response by modulating ICE1 protein stability. Dev. Cell 43 , 618–629 e5 (2017).
Danquah, A. et al. Identification and characterization of an ABA-activated MAP kinase cascade in Arabidopsis thaliana . Plant J. 82 , 232–244 (2015).
de Zelicourt, A., Colcombet, J. & Hirt, H. The role of MAPK modules and ABA during abiotic stress signaling. Trends Plant Sci. 21 , 677–685 (2016).
Qi, J. et al. Reactive oxygen species signaling and stomatal movement in plant responses to drought stress and pathogen attack. J. Integr. Plant Biol. 60 , 805–826 (2018).
Chan, K. X., Phua, S. Y., Crisp, P., McQuinn, R. & Pogson, B. J. Learning the languages of the chloroplast: retrograde signaling and beyond. Annu. Rev. Plant Biol. 67 , 25–53 (2016).
Pesaresi, P. & Kim, C. Current understanding of GUN1: a key mediator involved in biogenic retrograde signaling. Plant Cell Rep. 38 , 819–823 (2019).
Hua, D. P. et al. A plasma membrane receptor kinase, GHR1, mediates abscisic acid- and hydrogen peroxide-regulated stomatal movement in Arabidopsis . Plant Cell 24 , 2546–2561 (2012).
Wu, F. H. et al. Hydrogen peroxide sensor HPCA1 is an LRR receptor kinase in Arabidopsis . Nature 578 , 577–581 (2020).
Brandt, B. et al. Calcium specificity signaling mechanisms in abscisic acid signal transduction in Arabidopsis guard cells. eLife 4 , e03599 (2015).
PubMed Central Google Scholar
Geiger, D. et al. Guard cell anion channel SLAC1 is regulated by CDPK protein kinases with distinct Ca 2+ affinities. Proc. Natl Acad. Sci. USA 107 , 8023–8028 (2010).
Meinhard, M., Rodriguez, P. L. & Grill, E. The sensitivity of ABI2 to hydrogen peroxide links the abscisic acid-response regulator to redox signalling. Planta 214 , 775–782 (2002).
Miller, G. et al. The plant NADPH oxidase RBOHD mediates rapid systemic signaling in response to diverse stimuli. Sci. Signal. 2 , ra45 (2009).
Suzuki, N. et al. Temporal–spatial interaction between reactive oxygen species and abscisic acid regulates rapid systemic acclimation in plants. Plant Cell 25 , 3553–3569 (2013).
Choi, W. G. et al. Orchestrating rapid long-distance signaling in plants with Ca 2+ , ROS and electrical signals. Plant J. 90 , 698–707 (2017).
Choi, W. G., Toyota, M., Kim, S. H., Hilleary, R. & Gilroy, S. Salt stress-induced Ca 2+ waves are associated with rapid, long-distance root-to-shoot signaling in plants. Proc. Natl Acad. Sci. USA 111 , 6497–6502 (2014).
Zandalinas, S. I., Fichman, Y. & Mittler, R. Vascular bundles mediate systemic reactive oxygen signaling during light stress. Plant Cell 32 , 3425–3435 (2020).
Fichman, Y., Myers, R. J., Grant, D. G. & Mittler, R. Plasmodesmata-localized proteins and ROS orchestrate light-induced rapid systemic signaling in Arabidopsis . Sci. Signal. 14 , eabf0322 (2021).
Zandalinas, S. I. & Mittler, R. Vascular and non-vascular transmission of systemic reactive oxygen signals during wounding and heat stress. Plant Physiol. 186 , 1721–1733 (2021).
Takahashi, F. et al. A small peptide modulates stomatal control via abscisic acid in long-distance signalling. Nature 556 , 235–238 (2018). This work demonstrates small peptide-mediated root to shoot signalling in response to drought .
Dinneny, J. R. Developmental responses to water and salinity in root systems. Annu. Rev. Cell Dev. Biol. 35 , 239–257 (2019).
Leftley, N., Banda, J., Pandey, B., Bennett, M. & Voss, U. Uncovering how auxin optimizes root systems architecture in response to environmental stresses. Cold Spring Harb. Perspect. Biol . https://doi.org/10.1101/cshperspect.a040014 (2021).
Article PubMed Google Scholar
Hua, J. From freezing to scorching, transcriptional responses to temperature variations in plants. Curr. Opin. Plant Biol. 12 , 568–573 (2009).
Jacob, P., Hirt, H. & Bendahmane, A. The heat-shock protein/chaperone network and multiple stress resistance. Plant Biotechnol. J. 15 , 405–414 (2017).
Ma, S. & Bohnert, H. J. Integration of Arabidopsis thaliana stress-related transcript profiles, promoter structures, and cell-specific expression. Genome Biol. 8 , R49 (2007). This study in A. thaliana summarizes common transcriptional responses to various environmental stresses .
Zhu, J. K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 53 , 247–273 (2002).
Narusaka, Y. et al. Interaction between two cis -acting elements, ABRE and DRE, in ABA-dependent expression of Arabidopsis rd29A gene in response to dehydration and high-salinity stresses. Plant J. 34 , 137–148 (2003).
Yamaguchi-Shinozaki, K. & Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol. 57 , 781–803 (2006).
Liu, Q. et al. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis . Plant Cell 10 , 1391–1406 (1998).
Stockinger, E. J., Gilmour, S. J. & Thomashow, M. F. Arabidopsis thaliana CBF1 encodes an AP2 domain-containing transcriptional activator that binds to the C-repeat/DRE, a cis -acting DNA regulatory element that stimulates transcription in response to low temperature and water deficit. Proc. Natl Acad. Sci. USA 94 , 1035–1040 (1997).
Shinwari, Z. K. et al. An Arabidopsis gene family encoding DRE/CRT binding proteins involved in low-temperature-responsive gene expression. Biochem. Biophys. Res. Commun. 250 , 161–170 (1998).
Haake, V. et al. Transcription factor CBF4 is a regulator of drought adaptation in Arabidopsis . Plant Physiol. 130 , 639–648 (2002).
Nakashima, K. et al. Organization and expression of two Arabidopsis DREB2 genes encoding DRE-binding proteins involved in dehydration- and high-salinity-responsive gene expression. Plant Mol. Biol. 42 , 657–665 (2000).
Fowler, S. & Thomashow, M. F. Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway. Plant Cell 14 , 1675–1690 (2002).
Kreps, J. A. et al. Transcriptome changes for Arabidopsis in response to salt, osmotic, and cold stress. Plant Physiol. 130 , 2129–2141 (2002).
Seki, M. et al. Monitoring the expression pattern of around 7,000 Arabidopsis genes under ABA treatments using a full-length cDNA microarray. Funct. Integr.Genomics 2 , 282–291 (2002).
Chinnusamy, V., Zhu, J. & Zhu, J. K. Cold stress regulation of gene expression in plants. Trends Plant Sci. 12 , 444–451 (2007).
Tang, K. et al. The transcription factor ICE1 functions in cold stress response by binding to the promoters of CBF and COR genes. J. Integr. Plant Biol. 62 , 258–263 (2020).
Thomashow, M. F. Plant cold acclimation: freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Physiol. Plant Mol. Biol. 50 , 571–599 (1999).
Medina, J., Catalá, R. & Salinas, J. The CBFs: three Arabidopsis transcription factors to cold acclimate. Plant Sci. 180 , 3–11 (2011).
Santiago, J. et al. Modulation of drought resistance by the abscisic acid receptor PYL5 through inhibition of clade A PP2Cs. Plant J. 60 , 575–588 (2009).
Szostkiewicz, I. et al. Closely related receptor complexes differ in their ABA selectivity and sensitivity. Plant J. 61 , 25–35 (2010).
Claeys, H., Van Landeghem, S., Dubois, M., Maleux, K. & Inze, D. What is stress? Dose–response effects in commonly used in vitro stress assays. Plant Physiol. 165 , 519–527 (2014). This report highlights that plant abiotic stress responses are strongly dependent on the stress levels .
Watkinson, J. I. et al. Photosynthetic acclimation is reflected in specific patterns of gene expression in drought-stressed loblolly pine. Plant Physiol. 133 , 1702–1716 (2003).
Hugouvieux, V., Kwak, J. M. & Schroeder, J. I. An mRNA cap binding protein, ABH1, modulates early abscisic acid signal transduction in Arabidopsis . Cell 106 , 477–487 (2001).
Okamoto, M. et al. Sm-like protein-mediated RNA metabolism is required for heat stress tolerance in arabidopsis. Front. Plant Sci. 7 , 1079 (2016).
Wu, S. J., Wang, L. C., Yeh, C. H., Lu, C. A. & Wu, S. J. Isolation and characterization of the Arabidopsis heat-intolerant 2 (hit2) mutant reveal the essential role of the nuclear export receptor EXPORTIN1A (XPO1A) in plant heat tolerance. N. Phytol. 186 , 833–842 (2010).
Zhan, X. et al. An Arabidopsis PWI and RRM motif-containing protein is critical for pre-mRNA splicing and ABA responses. Nat. Commun. 6 , 8139 (2015).
Gong, Z. et al. RNA helicase-like protein as an early regulator of transcription factors for plant chilling and freezing tolerance. Proc. Natl Acad. Sci. USA 99 , 11507–11512 (2002).
Guan, Q. et al. A DEAD box RNA helicase is critical for pre-mRNA splicing, cold-responsive gene regulation, and cold tolerance in Arabidopsis . Plant Cell 25 , 342–356 (2013).
Lu, C. A. et al. DEAD-Box RNA helicase 42 plays a critical role in pre-mRNA splicing under cold stress. Plant Physiol. 182 , 255–271 (2020).
Wang, B. et al. The DEAD-box RNA helicase SHI2 functions in repression of salt-inducible genes and regulation of cold-inducible gene splicing. J. Exp. Bot. 71 , 1598–1613 (2020).
Ling, Y. et al. Pre-mRNA splicing repression triggers abiotic stress signaling in plants. Plant J. 89 , 291–309 (2017).
Marquez, Y., Brown, J. W., Simpson, C., Barta, A. & Kalyna, M. Transcriptome survey reveals increased complexity of the alternative splicing landscape in Arabidopsis . Genome Res. 22 , 1184–1195 (2012).
Ding, F. et al. Genome-wide analysis of alternative splicing of pre-mRNA under salt stress in Arabidopsis . BMC Genomics 15 , 431 (2014).
Li, Y. et al. Comprehensive profiling of alternative splicing landscape during cold acclimation in tea plant. BMC Genomics 21 , 65 (2020).
Wang, Z. J. et al. ABA signalling is fine-tuned by antagonistic HAB1 variants. Nat. Commun. 6 , 8138 (2015).
Gu, J. et al. Spliceosomal protein U1A is involved in alternative splicing and salt stress tolerance in Arabidopsis thaliana . Nucleic Acids Res. 46 , 1777–1792 (2018).
Chong, G. L., Foo, M. H., Lin, W. D., Wong, M. M. & Verslues, P. E. Highly ABA-induced 1 (HAI1)-interacting protein HIN1 and drought acclimation-enhanced splicing efficiency at intron retention sites. Proc. Natl Acad. Sci. USA 116 , 22376–22385 (2019).
Chakrabarti, M., de Lorenzo, L., Abdel-Ghany, S. E., Reddy, A. S. N. & Hunt, A. G. Wide-ranging transcriptome remodelling mediated by alternative polyadenylation in response to abiotic stresses in Sorghum. Plant J. 102 , 916–930 (2020).
Tellez-Robledo, B. et al. The polyadenylation factor FIP1 is important for plant development and root responses to abiotic stresses. Plant J. 99 , 1203–1219 (2019).
Ye, C. T., Zhou, Q., Wu, X. H., Ji, G. L. & Li, Q. S. Q. Genome-wide alternative polyadenylation dynamics in response to biotic and abiotic stresses in rice. Ecotoxicol. Environ. Saf. 183 , 109485 (2019).
Rogers, K. & Chen, X. Biogenesis, turnover, and mode of action of plant microRNAs. Plant Cell 25 , 2383–2399 (2013).
Sunkar, R., Chinnusamy, V., Zhu, J. & Zhu, J. K. Small RNAs as big players in plant abiotic stress responses and nutrient deprivation. Trends Plant Sci. 12 , 301–309 (2007).
Sunkar, R., Kapoor, A. & Zhu, J. K. Posttranscriptional induction of two Cu/Zn superoxide dismutase genes in Arabidopsis is mediated by downregulation of miR398 and important for oxidative stress tolerance. Plant Cell 18 , 2051–2065 (2006).
Liu, Y. et al. MiR319 mediated salt tolerance by ethylene. Plant Biotechnol. J. 17 , 2370–2383 (2019).
Zhou, M. et al. Constitutive expression of a miR319 gene alters plant development and enhances salt and drought tolerance in transgenic creeping bentgrass. Plant Physiol. 161 , 1375–1391 (2013).
Yang, C. H. et al. Overexpression of microRNA319 impacts leaf morphogenesis and leads to enhanced cold tolerance in rice ( Oryza sativa L.). Plant Cell Env. 36 , 2207–2218 (2013).
Merret, R. et al. Heat-induced ribosome pausing triggers mRNA co-translational decay in Arabidopsis thaliana . Nucleic Acids Res. 43 , 4121–4132 (2015). This work reveals that heat-induced ribosomal pausing in A. thaliana preferentially occurs on transcripts coding for HSC/HSP70 chaperone targets .
Merret, R. et al. XRN4 and LARP1 are required for a heat-triggered mRNA decay pathway involved in plant acclimation and survival during thermal stress. Cell Rep. 5 , 1279–1293 (2013).
Nguyen, A. H. et al. Loss of Arabidopsis 5′–3′ exoribonuclease AtXRN4 function enhances heat stress tolerance of plants subjected to severe heat stress. Plant Cell Physiol. 56 , 1762–1772 (2015).
Merret, R. et al. Heat shock protein HSP101 affects the release of ribosomal protein mRNAs for recovery after heat shock. Plant Physiol. 174 , 1216–1225 (2017).
Zhang, L. et al. Mutations in eIF5B confer thermosensitive and pleiotropic phenotypes via translation defects in Arabidopsis thaliana . Plant Cell 29 , 1952–1969 (2017).
Yu, H. et al. STCH4/REIL2 confers cold stress tolerance in Arabidopsis by promoting rRNA processing and CBF protein translation. Cell Rep. 30 , 229–242.e5 (2020).
Wang, S. et al. Chloroplast RNA-binding protein RBD1 promotes chilling tolerance through 23S rRNA processing in Arabidopsis . PLoS Genet. 12 , e1006027 (2016).
Ding, Y. et al. EGR2 phosphatase regulates OST1 kinase activity and freezing tolerance in Arabidopsis . EMBO J. 38 , e99819 (2019). This study reports a mechanism for ABA-independent activation of SnRK2.6 in response to cold stress .
Kilian, J. et al. The AtGenExpress global stress expression data set: protocols, evaluation and model data analysis of UV-B light, drought and cold stress responses. Plant J. 50 , 347–363 (2007).
Willems, P. et al. The Plant PTM Viewer, a central resource for exploring plant protein modifications. Plant J. 99 , 752–762 (2019).
Feng, J., Chen, L. & Zuo, J. Protein S-nitrosylation in plants: current progresses and challenges. J. Integr. Plant Biol. 61 , 1206–1223 (2019).
Matamoros, M. A. & Becana, M. Molecular responses of legumes to abiotic stress: protein post-translational modifications and redox signaling. J. Exp. Bot. 72 , 5876–5892 (2021).
Hu, J. et al. Nitric oxide regulates protein methylation during stress responses in plants. Mol. Cell 67 , 702–710.e4 (2017).
Zhang, H., Lang, Z. & Zhu, J. K. Dynamics and function of DNA methylation in plants. Nat. Rev. Mol. Cell Biol. 19 , 489–506 (2018).
Chang, Y. N. et al. Epigenetic regulation in plant abiotic stress responses. J. Integr. Plant Biol. 62 , 563–580 (2020).
Chinnusamy, V., Dalal, M. & Zhu, J. K. Epigenetic regulation of abiotic stress responses in plants. Plant Abiotic Stress https://doi.org/10.1002/9781118764374.ch8 (2014).
Article Google Scholar
Khan, A. R. et al. Vernalization treatment induces site-specific DNA hypermethylation at the VERNALIZATION-A1 (VRN-A1) locus in hexaploid winter wheat. BMC Plant Biol. 13 , 209 (2013).
Xu, R. et al. Salt-induced transcription factor MYB74 is regulated by the RNA-directed DNA methylation pathway in Arabidopsis . J. Exp. Bot. 66 , 5997–6008 (2015).
Yong-Villalobos, L. et al. Methylome analysis reveals an important role for epigenetic changes in the regulation of the Arabidopsis response to phosphate starvation. Proc. Natl Acad. Sci. USA 112 , E7293–E7302 (2015).
Baek, D. et al. Regulated AtHKT1 gene expression by a distal enhancer element and DNA methylation in the promoter plays an important role in salt tolerance. Plant Cell Physiol. 52 , 149–161 (2011).
Zheng, M. et al. Histone acetyltransferase GCN5 contributes to cell wall integrity and salt stress tolerance by altering the expression of cellulose synthesis genes. Plant J. 97 , 587–602 (2019).
Zhu, Y. et al. The arabidopsis nodulin homeobox factor AtNDX interacts with AtRING1A/B and negatively regulates abscisic acid signaling. Plant Cell 32 , 703–721 (2020).
Zhang, B. et al. Chilling-induced tomato flavor loss is associated with altered volatile synthesis and transient changes in DNA methylation. Proc. Natl Acad. Sci. USA 113 , 12580–12585 (2016).
Zemach, A. et al. The Arabidopsis nucleosome remodeler DDM1 allows DNA methyltransferases to access H1-containing heterochromatin. Cell 153 , 193–205 (2013).
Vaillant, I., Schubert, I., Tourmente, S. & Mathieu, O. MOM1 mediates DNA-methylation-independent silencing of repetitive sequences in Arabidopsis . EMBO Rep. 7 , 1273–1278 (2006).
Iwasaki, M. & Paszkowski, J. Identification of genes preventing transgenerational transmission of stress-induced epigenetic states. Proc. Natl Acad. Sci. USA 111 , 8547–8552 (2014). This study identifies MOM1 and DDM1 as two key factors that prevent transgenerational transmission of stress-induced epigenetic states .
Jiang, C. et al. Environmentally responsive genome-wide accumulation of de novo Arabidopsis thaliana mutations and epimutations. Genome Res. 24 , 1821–1829 (2014).
Lang-Mladek, C. et al. Transgenerational inheritance and resetting of stress-induced loss of epigenetic gene silencing in Arabidopsis . Mol. Plant 3 , 594–602 (2010).
Wibowo, A. et al. Hyperosmotic stress memory in Arabidopsis is mediated by distinct epigenetically labile sites in the genome and is restricted in the male germline by DNA glycosylase activity. eLife 5 , e13546 (2016).
Sanchez, D. H. & Paszkowski, J. Heat-induced release of epigenetic silencing reveals the concealed role of an imprinted plant gene. PLoS Genet. 10 , e1004806 (2014).
Hirsch, C. N. et al. Insights into the maize pan-genome and pan-transcriptome. Plant Cell 26 , 121–135 (2014).
Liu, Y. C. et al. Pan-genome of wild and cultivated soybeans. Cell 182 , 162–176.e13 (2020).
Qin, P. et al. Pan-genome analysis of 33 genetically diverse rice accessions reveals hidden genomic variations. Cell 184 , 3542–3558.e16 (2021).
Munns, R. et al. Wheat grain yield on saline soils is improved by an ancestral Na + transporter gene. Nat. Biotechnol. 30 , 360-U173 (2012).
Ren, Z. H. et al. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Genet. 37 , 1141–1146 (2005).
Zhang, M. et al. A retrotransposon in an HKT1 family sodium transporter causes variation of leaf Na + exclusion and salt tolerance in maize. N. Phytol. 217 , 1161–1176 (2018).
Li, X. M. et al. Natural alleles of a proteasome α2 subunit gene contribute to thermotolerance and adaptation of African rice. Nat. Genet. 47 , 827–833 (2015).
Wang, Z. et al. Loss of salt tolerance during tomato domestication conferred by variation in a Na + /K + transporter. EMBO J. 39 , e103256 (2020).
Wang, Z. et al. Natural variations in SlSOS1 contribute to the loss of salt tolerance during tomato domestication. Plant Biotechnol. J. 19 , 20–22 (2021).
Mao, H. D. et al. A transposable element in a NAC gene is associated with drought tolerance in maize seedlings. Nat. Commun. 6 , 8326 (2015).
Wang, X. et al. Genetic variation in ZmVPP1 contributes to drought tolerance in maize seedlings. Nat. Genet. 48 , 1233–1241 (2016).
Cui, M. et al. Induced over-expression of the transcription factor OsDREB2A improves drought tolerance in rice. Plant Physiol. Biochem. 49 , 1384–1391 (2011).
Mallikarjuna, G., Mallikarjuna, K., Reddy, M. K. & Kaul, T. Expression of OsDREB2A transcription factor confers enhanced dehydration and salt stress tolerance in rice ( Oryza sativa L.). Biotechnol. Lett. 33 , 1689–1697 (2011).
Gupta, B. K. et al. Manipulation of glyoxalase pathway confers tolerance to multiple stresses in rice. Plant Cell Env. 41 , 1186–1200 (2018).
Zhang, J. et al. Knockdown of rice microRNA166 confers drought resistance by causing leaf rolling and altering stem xylem development. Plant Physiol. 176 , 2082–2094 (2018).
Waltz, E. Beating the heat. Nat. Biotechnol. 32 , 610–613 (2014).
Simmons, C. R. et al. Successes and insights of an industry biotech program to enhance maize agronomic traits. Plant Sci. 307 , 110899 (2021).
Lou, D., Wang, H. & Yu, D. The sucrose non-fermenting-1-related protein kinases SAPK1 and SAPK2 function collaboratively as positive regulators of salt stress tolerance in rice. BMC Plant Biol. 18 , 203 (2018).
Lu, Y. et al. Targeted, efficient sequence insertion and replacement in rice. Nat. Biotechnol. 38 , 1402–1407 (2020). This paper reports a technology that allows highly efficient insertion of transcriptional or translational regulatory sequences in any gene of interest .
Wang, M. G. et al. Optimizing base editors for improved efficiency and expanded editing scope in rice. Plant Biotechnol. J. 17 , 1697–1699 (2019).
Zhan, X., Lu, Y., Zhu, J. K. & Botella, J. R. Genome editing for plant research and crop improvement. J. Integr. Plant Biol. 63 , 3–33 (2020).
Zhu, H., Li, C. & Gao, C. Applications of CRISPR–Cas in agriculture and plant biotechnology. Nat. Rev. Mol. Cell Biol. 21 , 661–677 (2020).
Morran, S. et al. Improvement of stress tolerance of wheat and barley by modulation of expression of DREB/CBF factors. Plant Biotechnol. J. 9 , 230–249 (2011).
Nakashima, K. et al. Comparative functional analysis of six drought-responsive promoters in transgenic rice. Planta 239 , 47–60 (2014).
Wang, P. C. et al. Reciprocal regulation of the TOR kinase and ABA receptor balances plant growth and stress response. Mol. Cell 69 , 100–112 (2018). This work elucidates an important mechanism underlying the antagonism between stress response and plant growth .
Dobrenel, T. et al. TOR signaling and nutrient sensing. Annu. Rev. Plant Biol. 67 , 261–285 (2016).
Cai, Z. Y. et al. GSK3-like kinases positively modulate abscisic acid signaling through phosphorylating subgroup III SnRK2s in Arabidopsis . Proc. Natl Acad. Sci. USA 111 , 9651–9656 (2014).
Li, J. F. et al. The GSK3-like kinase BIN2 is a molecular switch between the salt stress response and growth recovery in Arabidopsis thaliana . Dev. Cell 55 , 367–380 (2020).
Nolan, T. M., Vukasinovic, N., Liu, D. R., Russinova, E. & Yin, Y. H. Brassinosteroids: multidimensional regulators of plant growth, development, and stress responses. Plant Cell 32 , 295–318 (2020).
Srivastava, M. et al. SUMO conjugation to BZR1 enables brassinosteroid signaling to integrate environmental cues to shape plant growth. Curr. Biol. 30 , 1410–1423 (2020).
Wang, H. J. et al. Abscisic acid signaling inhibits brassinosteroid signaling through dampening the dephosphorylation of BIN2 by ABI1 and ABI2. Mol. Plant 11 , 315–325 (2018).
Cao, M. J. et al. An ABA-mimicking ligand that reduces water loss and promotes drought resistance in plants. Cell Res. 23 , 1043–1054 (2013).
Singh, R., Bhardwaj, V. K., Sharma, J. & Purohit, R. Identification of novel and selective agonists for ABA receptor PYL3. Plant Physiol. Biochem. 154 , 387–395 (2020).
Vaidya, A. S. et al. Dynamic control of plant water use using designed ABA receptor agonists. Science 366 , eaaw8848 (2019).
Cao, M. J. et al. Combining chemical and genetic approaches to increase drought resistance in plants. Nat. Commun. 8 , 1183 (2017). This report demonstrates the power of combining genetic and chemical approaches for improving plant drought stress resistance .
Reinhold-Hurek, B., Bunger, W., Burbano, C. S., Sabale, M. & Hurek, T. Roots shaping their microbiome: global hotspots for microbial activity. Annu. Rev. Phytopathol. 53 , 403–424 (2015).
Vilchez, J. I. et al. DNA demethylases are required for myo-inositol-mediated mutualism between plants and beneficial rhizobacteria. Nat. Plants 6 , 983–995 (2020).
Liu, X. M. & Zhang, H. The effects of bacterial volatile emissions on plant abiotic stress tolerance. Front. Plant Sci. 6 , 774 (2015).
Lugtenberg, B. & Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 63 , 541–556 (2009).
Chen, K. et al. BONZAI proteins control global osmotic stress responses in plants. Curr. Biol. 30 , 4815–4825.e4 (2020).
Li, H. et al. MPK3-and MPK6-mediated ICE1 phosphorylation negatively regulates ICE1 stability and freezing tolerance in Arabidopsis . Dev. Cell 43 , 630–642 (2017).
Ding, Y. et al. OST1-mediated BTF3L phosphorylation positively regulates CBFs during plant cold responses. EMBO J. 37 , e98228 (2018).
Wang, X. et al. PUB25 and PUB26 promote plant freezing tolerance by degrading the cold signaling negative regulator MYB15. Dev. Cell 51 , 222–235.e5 (2019).
Liu, Z. et al. Plasma membrane CRPK1-mediated phosphorylation of 14-3-3 proteins induces their nuclear import to fine-tune CBF signaling during cold response. Mol. Cell 66 , 117–128.e5 (2017).
Download references
Acknowledgements
The authors apologize to those colleagues whose work is not cited owing to space constraints. H.Z. and J-K.Z. have been supported by the Chinese Academy of Sciences. Z.G. acknowledges grants from the National Science Foundation of China (31730007, 32030008, 31921001).
Author information
Authors and affiliations.
Shanghai Center for Plant Stress Biology, Center for Excellence in Molecular Plant Sciences, Chinese Academy of Sciences, Shanghai, China
Huiming Zhang & Jian-Kang Zhu
Department of Plant Science and Landscape Architecture, University of Maryland, College Park, MD, USA
Jianhua Zhu
State Key Laboratory of Plant Physiology and Biochemistry, College of Biological Sciences, China Agricultural University, Beijing, China
Zhizhong Gong
Institute of Life Science and Green Development, School of Life Sciences, Hebei University, Baoding, China
You can also search for this author in PubMed Google Scholar
Contributions
J.-K. Z. and H.Z. researched, discussed and wrote the article. All authors reviewed and/or edited the article before submission.
Corresponding authors
Correspondence to Huiming Zhang or Jian-Kang Zhu .
Ethics declarations
Competing interests.
The authors declare no competing interests.
Additional information
Peer review information.
Nature Reviews Genetics thanks F. Van Breusegem and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Increased extracellular osmolarity that leads to cell dehydration.
Acidic polysaccharides that are critical for plant cell wall properties such as cohesion and electrostatic potential.
Hydroxyproline-rich glycoproteins that are essential components of the cell wall.
A qualitative change in the state of a system under a continuous change in an external parameter, such as the reversible condensation of certain proteins within cells in response to changing temperature.
Signalling from organelles to the nucleus.
A systemic response to abiotic stimuli, which involves long-distance communication among cells belonging to different tissues or organs.
Isogenic alleles with different epigenetic modifications that are passed from generation to generation.
Chromosome regions that statistically contribute to the variability of a quantitative phenotype.
Approaches searching the entire genome using single-nucleotide polymorphisms for chromosome regions that show consistent correlation with a particular phenotype, within a large population of individuals with contrasting and varying degree of the phenotype.
An approach for breeding in which the selection of desired individuals is based on the score of molecular markers.
The soil layer that surrounds, and is influenced by, plant roots.
All microorganisms that inhabit a defined region.
Rights and permissions
Reprints and permissions
About this article
Cite this article.
Zhang, H., Zhu, J., Gong, Z. et al. Abiotic stress responses in plants. Nat Rev Genet 23 , 104–119 (2022). https://doi.org/10.1038/s41576-021-00413-0
Download citation
Accepted : 17 August 2021
Published : 24 September 2021
Issue Date : February 2022
DOI : https://doi.org/10.1038/s41576-021-00413-0
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
This article is cited by
Genome-wide analysis and prediction of chloroplast and mitochondrial rna editing sites of agc gene family in cotton (gossypium hirsutum l.) for abiotic stress tolerance.
- Furqan Ahmad
- Muhammad Abdullah
- Ehab A. A. Salama
BMC Plant Biology (2024)
Genome-wide identification and analysis of abiotic stress responsiveness of the mitogen-activated protein kinase gene family in Medicago sativa L.
- Xianyang Li
An integrated physiological indicator and transcriptomic analysis reveals the response of soybean buds to high-temperature stress
- Xiaobo Wang
Comprehensive identification of maize ZmE2F transcription factors and the positive role of ZmE2F6 in response to drought stress
- Haoqiang Yu
BMC Genomics (2024)
Genome-wide analysis of the WRKY gene family and their response to low-temperature stress in elephant grass
- Linkai Huang
Quick links
- Explore articles by subject
- Guide to authors
- Editorial policies
Sign up for the Nature Briefing newsletter — what matters in science, free to your inbox daily.

EDITORIAL article
Editorial: abiotic stress in plants: sustainability and productivity.

- 1 Faculty of Agricultural Sciences, Federal University of Grande Dourados, Dourados, Mato Grosso do Sul, Brazil
- 2 Department of Agriculture, Mediterranea University of Reggio, Calabria, Italy
- 3 Centre of Natural and Exact Sciences, Federal University of Santa Maria, Santa Maria, Rio Grande do Sul, Brazil
Editorial on the Research Topic Abiotic stress in plants: sustainability and productivity
The Research Topic Abiotic Stress in Plants: Sustainability and Productivity was dedicated to study the effects of stressful conditions of abiotic origin, such as water and thermal stress, heavy metals and soil salinity, on crops and tree species, with focus on molecular and physio metabolic responses or adjustments. Innovative management and cultural treatments as a promising practice to induce tolerance and favor plant survival, as well as efficient recovery and productivity, were also considered in the thirteen articles published on this topic.
In their review, Yang et al. discuss recent advances in heat perception mechanisms in plants, with focus on second messengers. Furthermore, this review reported the regulatory mechanisms that involve specific transcription factors. Calcium ion, hydrogen peroxide, and nitric oxide have emerged as key players in heat perception. After discussing the roles of transcription, thermotolerance, and temperature targeting factors, such as plasma membrane-associated thermosensors, the authors paid attention to unsolved questions in the field of heat perception that require further investigation in the future.
The review by Qin et al . reports the potential of porous fiber materials (PFM) made from mineral rocks in conserving soil and water in areas where flooding and drought may occur. The effectiveness of PFM depends on their porosity and permeability, which, if high they can increase the capacity to retain, replace and infiltrate water into the soil during rainfalls, thereby reducing the risk of flooding, runoff and nutrients loss.
In two of the papers presented here, the issue of abiotic stress effects modulated by the plant’s developmental stage is examined. Zhang et al. observe that, at any stage, mild water deficit reduces the crop coefficient and evapotranspiration intensity in watermelon, and the water demand differs among the stages, increasing rapidly from the seedling stage to vines.
Similarly, Chen et al. describe that management of deficit irrigation is a strategy to conserve water in agriculture. In their study with sunflower, they also report that mild water deficit, both at the seedling stage and maturity, promoted increase in water use efficiency by leaves. The authors conclude that mild water deficit may be an effective irrigation strategy for sunflower production in the cold and arid environment of Northwest China.
In addition to the above focus on developmental stages, the research reported here shows that cultivars and genotypes of the same species respond differently to abiotic stresses as well as the resources used to mitigate adverse environmental conditions. In this context, Safarpour et al. report that, although considered as resistant plant species, barley plants exposed to water stress showed decreases in growth and grain production, as well as an enhanced antioxidants' response. The barley cultivar showing the highest proline content and catalase activity was the one with the highest biomass production and grain yield, which led to the conclusion that foliar urea applications can be effective to increase water stress tolerance, by improving the plant physiological performance.
Zhou et al. evaluated the effect of irrigation management strategies associated with nitrogen fertilization on eggplant, and verified that mild water deficit combined with medium nitrogen application rate (W1N2) is the right choice for growing conditions in arid environment, providing better water and nitrogen use efficiency and productivity.
The germplasm variability in stress responses was also studied by Li et al. , who identified drought-resistant genotypes with potential value for breeding programs within a collection of 42 lettuce genotypes. Drought-resistance was found to be associated with little increases in stomata density, the production of superoxide and the content of malondialdehyde (MDA), but also with large increase in the activities of antioxidant enzymes, on the other hand.
In the same vein, Kumar et al. with the aim of identifying wheat genotypes tolerant to heat stress, verified that those tolerant ones maintained balanced phenological-physiobiochemical characteristics and high activities of their antioxidant enzymes. They also suggested that high photosynthesis and delayed senescence must be the best selection parameters for the heat tolerance in wheat.
The tolerance to a single stress or the combination of heat and drought stresses must be variable according to the cotton genotype, presenting different responses for enzymatic activity and non-enzymatic compounds ( Zafar et al. ). According to these authors, under combined stress, four genotypes exhibited superior performance in terms of agronomic traits and fiber quality, while others maintained gas exchange and relative water content, decreased their levels of H 2 O 2 and MDA and increased levels of chlorophylls, carotenoids and activity of antioxidant enzymes.
Many papers reported here suggest that alternative cultural treatments can mitigate the stressful effects of the environment. In a study on the effect of partial replacement of chemical fertilizer (CF) by Trichoderma biofertilizer (TF) on wolfberry cultivation in saline lands, Yan et al. observed that replacement with 75% CF improves the N use efficiency and promotes higher photosynthetic rate, resulting in greater biomass and fruit production.
Seeking for the mitigation of water stress effects on ‘jatobazeiro’ seedlings ( Hymenaea courbaril L.), Reis et al. confirmed the positive effect of intermediate shading of 30 and 50%, which accelerated the recovery of the photosynthetic rate after the resumption of irrigation. As a consequence, the cultivation under water deficit without shading (0%) should not be adopted for seedlings production in this species.
Considering that heavy metals pollution reduces the yield and quality of vegetables, Sun et al. demonstrated that the foliar spraying with zinc oxide nanoparticles (ZnO NPs) on tomato seedlings improved Cd tolerance, increased photosynthesis efficiency and antioxidant capacity, and reduced Cd accumulation in roots and leaves. Metabolomic analysis showed that exposure to ZnO NPs mitigates Cd toxicity, with higher effect on leaves than roots, with reduction of oxidative damage.
Once more in tomato seedlings, Turan et al. evaluated the potential of growth-promoting rhizobacteria (PGPR) to alleviate the stressful effect of drought, and verified that applying 4 L ha -1 of a biostimulant containing PGPR not only mitigated detrimental effects of water stress on hormonal balance and growth characteristics, but also restored plant growth and improved soil organic matter and the soil contents of total N, P, Ca, and Cu.
Author contributions
SS: Writing – review & editing, Writing – original draft. CS: Writing – review & editing, Writing – original draft. MB: Writing – review & editing, Writing – original draft. LT: Writing – original draft.
Acknowledgments
The Guest Editors would like to thank all the authors who contributed to this Research Topic. We would like to thank all the funding bodies and companies that contributed in some way to the articles published in this Research Topic.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Keywords: antioxidant enzymes, photosynthetic metabolism, regulatory mechanisms, salinity, thermal stress, transcription factors, water stress
Citation: Scalon SdPQ, Santos CC, Badiani M and Tabaldi LA (2024) Editorial: Abiotic stress in plants: sustainability and productivity. Front. Plant Sci. 15:1386174. doi: 10.3389/fpls.2024.1386174
Received: 14 February 2024; Accepted: 12 March 2024; Published: 20 March 2024.
Edited and Reviewed by:
Copyright © 2024 Scalon, Santos, Badiani and Tabaldi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY) . The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Silvana de Paula Quintão Scalon, c2lsdmFuYXNjYWxvbkB1ZmdkLmVkdS5icg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
- Open access
- Published: 17 November 2011
Effects of abiotic stress on plants: a systems biology perspective
- Grant R Cramer 1 ,
- Kaoru Urano 2 ,
- Serge Delrot 3 ,
- Mario Pezzotti 4 &
- Kazuo Shinozaki 2
BMC Plant Biology volume 11 , Article number: 163 ( 2011 ) Cite this article
109k Accesses
820 Citations
15 Altmetric
Metrics details
The natural environment for plants is composed of a complex set of abiotic stresses and biotic stresses. Plant responses to these stresses are equally complex. Systems biology approaches facilitate a multi-targeted approach by allowing one to identify regulatory hubs in complex networks. Systems biology takes the molecular parts (transcripts, proteins and metabolites) of an organism and attempts to fit them into functional networks or models designed to describe and predict the dynamic activities of that organism in different environments. In this review, research progress in plant responses to abiotic stresses is summarized from the physiological level to the molecular level. New insights obtained from the integration of omics datasets are highlighted. Gaps in our knowledge are identified, providing additional focus areas for crop improvement research in the future.
Recent advances in biotechnology have dramatically changed our capabilities for gene discovery and functional genomics. For the first time, we can now obtain a holistic "snapshot" of a cell with transcript, protein and metabolite profiling. Such a "systems biology" approach allows for a deeper understanding of physiologically complex processes and cellular function [ 1 ]. New models can be formed from the plethora of data collected and lead to new hypotheses generated from those models.
Understanding the function of genes is a major challenge of the post-genomic era. While many of the functions of individual parts are unknown, their function can sometimes be inferred through association with other known parts, providing a better understanding of the biological system as a whole. High throughput omics technologies are facilitating the identification of new genes and gene function. In addition, network reconstructions at the genome-scale are key to quantifying and characterizing the genotype to phenotype relationships [ 2 ].
In this review, we summarize recent progress on systematic analyses of plant responses to abiotic stress to include transcriptomics, metabolomics, proteomics, and other integrated approaches. Due to space limitations, we try to emphasize important perspectives, especially from what systems biology and omics approaches have provided in recent research on environmental stresses.
Plant responses to the environment are complex
Plants are complex organisms. It is difficult to find an estimate of the total number of cells in a plant. Estimates of the number of cells in the adaxial epidermal layer and palisade mesophyll of a simple Arabidopsis leaf are approximately 27,000 and 57,000 cells, respectively [ 3 ]. Another estimate of the adaxial side of the epidermal layer of the 7 th leaf of Arabidopsis was close to 100,000 cells [ 4 ] per cm 2 of leaf area. An Arabidopsis plant can grow as large as 14 g fresh weight with a leaf area of 258 cm 2 (11 g fresh weight) [ 5 ]. Thus, we estimate that a single Arabidopsis plant could have approximately 100 million cells (range of 30 to 150 million cells assuming 2.4 to 11 million cells per g fresh weight). A one million Kg redwood tree could possibly have 70 trillion cells assuming a cell size 100 times larger than an Arabidopsis cell. Combine that with developmental changes, cell differentiation and interactions with the environment and it is easy to see that there are an infinite number of permutations to this complexity.
There is additional complexity within the cell with multiple organelles, interactions between nuclear, plastidial and mitochondrial genomes, and between cellular territories that behave like symplastically isolated domains that are able to exchange transcription factors controlling gene expression and developmental stages across the plasmodesmata. A typical plant cell has more than 30,000 genes and an unknown number of proteins, which can have more than 200 known post-translational modifications (PTMs). The molecular responses of cells (and plants) to their environment are extremely complex.
Environmental limits to crop production
In 1982, Boyer indicated that environmental factors may limit crop production by as much as 70% [ 6 ]. A 2007 FAO report stated that only 3.5% of the global land area is not affected by some environmental constraint (see Table three point seven in http://www.fao.org/docrep/010/a1075e/a1075e00.htm ). While it is difficult to get accurate estimates of the effects of abiotic stress on crop production (see different estimates in Table 1 ), it is evident that abiotic stress continues to have a significant impact on plants based upon the percentage of land area affected and the number of scientific publications directed at various abiotic stresses (Table 1 ). If anything the environmental impacts are even more significant today; yields of the "big 5" food crops are expected to decline in many areas in the future due to the continued reduction of arable land, reduction of water resources and increased global warming trends and climate change [ 7 ].
This growing concern is reflected in the increasing number of publications focused on abiotic stresses. For example, since the pivotal review of systems biology by Kitano in 2002 [ 1 ], the number of papers published on abiotic stress in plants using a systems biology approach has increased exponentially (Figure 1 ).

The number of publications per year related to systems biology and abiotic stress . Key words used in the search of PubMed included: plant, systems biology, and abiotic stress (including stress sub-terms; e.g. drought or water deficit or dehydration). *The number for the year 2011 was estimated by doubling the 6-month value.
Multiple factors limit plant growth
Fundamentally, plants require energy (light), water, carbon and mineral nutrients for growth. Abiotic stress is defined as environmental conditions that reduce growth and yield below optimum levels. Plant responses to abiotic stresses are dynamic and complex [ 8 , 9 ]; they are both elastic (reversible) and plastic (irreversible).
The plant responses to stress are dependent on the tissue or organ affected by the stress. For example, transcriptional responses to stress are tissue or cell specific in roots and are quite different depending on the stress involved [ 10 ]. In addition, the level and duration of stress (acute vs chronic) can have a significant effect on the complexity of the response [ 11 , 12 ].
Water deficit inhibits plant growth by reducing water uptake into the expanding cells, and alters enzymatically the rheological properties of the cell wall; for example, by the activity of ROS (reactive oxygen species) on cell wall enzymes [ 8 ]. In addition, water deficit alters the cell wall nonenzymatically; for example, by the interaction of pectate and calcium [ 13 ]. Furthermore, water conductance to the expanding cells is affected by aquaporin activity and xylem embolism [ 14 – 17 ]. The initial growth inhibition by water deficit occurs prior to any inhibition of photosynthesis or respiration [ 18 , 19 ].
The growth limitation is in part due to the fundamental nature of newly divided cells encasing the xylem in the growing zone [ 20 , 21 ]. These cells act as a resistance to water flow to the expanding cells in the epidermis making it necessary for the plant to develop a larger water potential gradient. Growth is limited by the plant's ability to osmotically adjust or conduct water. The epidermal cells can increase the water potential gradient by osmotic adjustment, which may be largely supplied by solutes from the phloem. Such solutes are supplied by photosynthesis that is also supplying energy for growth and other metabolic functions in the plant. With long-term stress, photosynthesis declines due to stomatal limitations for CO 2 uptake and increased photoinhibition from difficulties in dissipating excess light energy [ 12 ].
One of the earliest metabolic responses to abiotic stresses and the inhibition of growth is the inhibition of protein synthesis [ 22 – 25 ] and an increase in protein folding and processing [ 26 ]. Energy metabolism is affected as the stress becomes more severe (e.g. sugars, lipids and photosynthesis) [ 12 , 27 , 28 ]. Thus, there are gradual and complex changes in metabolism in response to stress.
Central regulators limit key plant processes
The plant molecular responses to abiotic stresses involve interactions and crosstalk with many molecular pathways [ 29 ]. Systems biology and omics approaches have been used to elucidate some of the key regulatory pathways in plant responses to abiotic stress.
One of the earliest signals in many abiotic stresses involve ROS and reactive nitrogen species (RNS), which modify enzyme activity and gene regulation [ 30 – 32 ]. ROS signaling in response to abiotic stresses and its interactions with hormones has been thoroughly reviewed [ 32 ]. ROS and RNS form a coordinated network that regulates many plant responses to the environment; there are a large number of studies on the oxidative effects of ROS on plant responses to abiotic stress, but only a few studies documenting the nitrosative effects of RNS [ 30 ].
Hormones are also important regulators of plant responses to abiotic stress (Figure 2 ). The two most important are abscisic acid (ABA) and ethylene [ 33 ]. ABA is a central regulator of many plant responses to environmental stresses, particularly osmotic stresses [ 9 , 34 – 36 ]. Its signaling can be very fast without involving transcriptional activity; a good example is the control of stomatal aperture by ABA through the biochemical regulation of ion and water transport processes [ 35 ]. There are slower responses to ABA involving transcriptional responses that regulate growth, germination and protective mechanisms.

A simplified working model of a signaling network of plant responses to abiotic stress . Ovals represent proteins, metabolites or processes. Metabolites have magenta color. Phosphorylated proteins have red circles with a P inside. Sumoylated protein has an orange circle with an S inside. The solid purple circle indicates that DREB2 needs modification to be activated. Solid lines represent direct connections; dotted lines represent indirect connections (acting through some intermediate molecule). The gray line indicates that this reaction has not been shown in plants. Not all linkages and details of stress and hormone effects are shown in this diagram in order to simplify the model. Abbreviations: ABA (abscisic acid), ANAC (Arabidopsis NAC domain-containing protein), CAMTA (calmodulin-binding transcription activator), CBL (calcineurin B-like interacting protein kinase), CCA (circadian clock associated), CPK (calcium-dependent protein kinase), DREB/CBF (dehydration response element binding protein/C-repeat binding factor), ETR1 (ethylene response 1), GCN2 (general control non-repressible 2), HSF (heat shock factor), ICE (inducer of CBF expression), MAPK (mitogen-activated protein kinase), LHY (late elongated hypocotyl), PA (phosphatidic acid), PP2C (protein phosphatase 2C), PRR (pseudo response regulator), PYR/PYL/RCAR (ABA receptors), RNS (reactive nitrogen species), ROS (reactive oxygen species), SIZ (SAP and Miz domain protein), SnRK (sucrose nonfermenting-1 related kinase), TFs (transcription factors), TOR (target of rapamycin), ZAT (zinc finger protein).
Recently, the essential components of ABA signaling have been identified, and their mode of action was clarified [ 37 ]. The current model of ABA signaling includes three core components, receptors (PYR/PYL/RCAR), protein phosphatases (PP2C) and protein kinases (SnRK2/OST1) [ 38 , 39 ]. The PYR/PYL/RCAR proteins were identified as soluble ABA receptors by two independent groups [ 38 , 39 ]. The 2C-type protein phosphatases (PP2C) including ABI1 and ABI2, were first identified from the ABA-insensitive Arabidopsis mutants abi1-1 and abi2-1 , and they act as global negative regulators of ABA signaling [ 40 ]. SNF1-related protein kinase 2 (SnRK2) is a family of protein kinases isolated as ABA-activated protein kinases [ 41 , 42 ]. In Arabidopsis, three members of this family, SRK2D/SnRK2.2, SRK2E/OST1/SnRK2.6, and SRK2I/SnRK2.3, regulate ABA signaling positively and globally, as shown in the triple knockout mutant srk2d srk2e srk2i ( srk2dei )/ snrk2.2 snrk2.3 snrk2.6 , which lacks ABA responses [ 43 ]. The PYR/PYL/RCAR - PP2C - SnRK2 complex plays a key role in ABA perception and signaling.
Studies of the transcriptional regulation of dehydration and salinity stresses have revealed both ABA-dependent and ABA-independent pathways [ 44 ]. Cellular dehydration under water limited conditions induces an increase in endogenous ABA levels that trigger downstream target genes encoding signaling factors, transcription factors, metabolic enzymes, and others [ 44 ]. In the vegetative stage, expression of ABA-responsive genes is mainly regulated by bZIP transcription factors (TFs) known as AREB/ABFs, which act in an ABA-responsive-element (ABRE) dependent manner [ 45 – 47 ]. Activation of ABA signaling cascades result in enhanced plant tolerance to dehydration stress. In contrast, a dehydration-responsive cis-acting element, DRE/CRT sequence and its DNA binding ERF/AP2-type TFs, DREB1/CBF and DREB2A, are related to the ABA-independent dehydration and temperature responsive pathways [ 44 ]. DREB1/CBFs function in cold-responsive gene expression [ 48 , 49 ], whereas DREB2s are involved in dehydration-responsive and heat-responsive gene expression [ 50 ].
Ethylene is also involved in many stress responses [ 51 – 53 ], including drought, ozone, flooding (hypoxia and anoxia), heat, chilling, wounding and UV-B light [ 31 , 33 , 53 ]. Ethylene signaling is well defined [ 51 , 52 ], and will not be discussed in detail here. There are known interactions between ethylene and ABA during drought [ 31 ], fruit ripening [ 54 , 55 ], and bud dormancy [ 56 ]. All of these interactions make the plant response to stress very complex [ 12 , 31 , 52 ].
In yeast, the well-documented central regulators of protein synthesis and energy are SnRK1 (Snf1/AMPK), TOR1 and GCN2 [ 57 – 60 ]. These proteins are largely controlled by the phosphorylation of enzymes; all three are protein kinases acting as key hubs in the coordination of metabolism during stressful conditions [ 61 ]. In plants, TOR activity is inhibited by osmotic stress and ABA [ 62 ] and GCN2 activity is stimulated by UV-light, amino acid starvation, ethylene, and cold stress [ 63 ]. SnRK1 responds to energy depletion, such as low light, nutrient deprivation or hypoxic conditions [ 64 , 65 ], and interacts with both glucose and ABA signaling pathways [ 66 ]. One of the results of this coordinated response is the inhibition of protein synthesis.
Many abiotic stresses directly or indirectly affect the synthesis, concentration, metabolism, transport and storage of sugars. Soluble sugars act as potential signals interacting with light, nitrogen and abiotic stress [ 67 – 69 ] to regulate plant growth and development; at least 10% of Arabidopsis genes are sugar-responsive [ 68 ]. Mutant analysis has revealed that sugar signaling interacts with ethylene [ 70 ], ABA [ 71 , 72 ], cytokinins [ 73 ], and light [ 74 , 75 ]. In grapevine, sugar and ABA signaling pathways interact to control sugar transport. An ASR (ABA, stress-, and ripening-induced) protein isolated from grape berries is upregulated synergistically by ABA and sugars, and upregulates the expression of a hexose transporter [ 76 ]. VVSK1, a GSK3 type protein kinase, is also induced by sugars and ABA, and upregulates the expression of several hexose transporters [ 77 ].
Stresses such as sugar starvation and lack of light stimulate SnRK1 activity ([ 64 ]. Suc-P synthase (SPS), 3-hydroxy-3-methylglutaryl-CoA reductase, nitrate reductase, and trehalose-6-P synthase are negatively regulated by SnRK1 phosphorylation [ 78 ], indicating that SnRK1 modulates metabolism by phosphorylating key metabolic enzymes. Post-translational redox modulation of ADPG-pyrophosphorylase, a key control of starch synthesis, by SnRK1 provides an interesting example of interactions between phosphorylation, redox control and sugar metabolism [ 79 ]. In Arabidopsis, SnRK1 kinase activity is itself increased by GRIK1 and GRIK2, which phosphorylate a threonine residue of the SnRK1 catalytic subunit [ 78 ]. SnRK2 interacts with ABA for the control of stomatal aperture and participates in the regulation of plant primary metabolism. Constitutive expression of SnRK2.6 drastically boosts sucrose and total soluble sugar levels in leaves, presumably by controlling SPS expression [ 80 ].
Systems biology approach to abiotic stress
In the post-genomic era, comprehensive analyses using three systematic approaches or omics have increased our understanding of the complex molecular regulatory networks associated with stress adaptation and tolerance. The first one is 'transcriptomics' for the analysis of coding and noncoding RNAs, and their expression profiles. The second one is 'metabolomics' that is a powerful tool to analyze a large number of metabolites. The third one is 'proteomics' in which protein and protein modification profiles offer an unprecedented understanding of regulatory networks. Protein complexes involved in signaling have been analyzed by a proteomics approach [ 81 , 82 ]. Integration of the different omics analyses facilitates abiotic stress signaling studies allowing for more robust identifications of molecular targets for future biotechnological applications in crops and trees.
Co-expression analyses identify regulatory hubs
An important application of transcriptomics data is co-expression analysis of target genes using on-line analytical tools, such as ATTED-II (reviewed by [ 83 ]). This approach is very promising for understanding gene-gene correlations and finding master genes in target conditions.
In a series of pioneering papers, Hirai et al. [ 84 , 85 ] identified MYB transcription factors regulating glucosinolate biosynthesis in Arabidopsis in response to S and N deficiency using an integrated transcriptomics and metabolomics approach. Genes and metabolites in glucosinolate metabolism were found to be coordinately regulated [ 84 ]. Co-expression analysis was used to identify two MYB transcription factors that positively regulate glucosinolate metabolism [ 85 ]. Then a knock out mutant and ectopic expression of one of the transcription factors was used to validate its positive role in glucosinolate metabolism. Previously unidentified genes were assigned to this biosynthetic pathway and a regulatory network model was constructed [ 85 ].
Mao et al. [ 86 ] performed a gene co-expression network analysis of 1094 microarrays of Arabidopsis using a non-targeted approach. They identified 382 modules in this network. The top three modules with the most nodes were: photosynthesis, response to oxidative stress and protein synthesis. Many of the modules also involved responses to environmental stresses. They constructed a cold-induced gene network from a subset of microarrays. The response to auxin stimulus was the most over-represented of the 18 significant modules.
Carrera et al. [ 87 ] used the InferGene application to construct a regulatory model of the Arabidopsis genome. They used datasets from 1,486 microarray experiments. Ten genes were predicted to be the most central regulatory hubs influencing the largest number of genes. Included in this set were transcription factor genes involved in auxin (KAN3), gibberellin (MYB29), abscisic acid (MYB121), ethylene (ERF1), and stress responses (ANAC036). They computed the top 12 gene subnetworks; four of these were related to biotic and abiotic stresses. Eighty-five percent of the predicted interactions of the 25% most connected transcription factors were validated in AtRegNet, the Arabidopsis thaliana Regulator Network http://arabidopsis.med.ohio-state.edu/moreNetwork.html .
Lorenz et al [ 88 ] investigated the drought response of loblolly pine roots and identified a number of hubs in the transcriptional network. Highly ranked hubs included thioredoxin, an inositol transporter, cardiolipin synthase/phosphatidyl transferase, 9-cis-expoxycarotenoid dioxygenase, zeatin O-glucosyltransferase and a SnRK2 kinase. These genes are involved in phospholipid metabolism, ABA biosynthesis and signaling, and cytokinin metabolism; they appear to be important in stress mediation.
Weston et al [ 89 ] used weighted co-expression analysis to define six modules for Arabidopsis responses to abiotic stress. Two hubs in the common response module were an ankyrin-repeat protein and genes involved in Ca signaling. They created a compendium of genomic signatures and linked them to their co-expression analysis. Using the same approach, they extended their analyses to the responses of three different plant species to heat and light [ 90 ]. Species-specific responses were found involving heat tolerance, heat-shock proteins, ROS, oligosaccharide metabolism and photosynthesis.
Time-series analyses reveal multiple phases in stress responses
Time-series analyses allow one to distinguish between primary and secondary responses to stress. In a comprehensive time-series transcriptomics analysis of 7 abiotic stresses on different Arabidopsis organs [ 28 ], a core set of genes (50% were transcription factors) of non-specific responses for all stresses were elucidated. Included in this set were the AZF2, ZAT10 and ZAT12 transcription factors. This initial response is thought to be involved in the readjustment of energy homeostasis in response to the stress. With time (after 1 h) more stress-specific profiles developed.
Sun et al [ 91 ] applied a complexity metric to a set of time series data of Arabidopsis with 9 different abiotic stresses. They found that genes with a higher complexity metric had longer 5' intergenic regions and a greater density of cis-regulatory motifs than the genes with a low complexity metric. Many of the cis-regulatory motifs identified were associated with previously characterized stress responses.
Vanderauwera et al. [ 92 ] investigated the effects of hydrogen peroxide (H 2 O 2 ) signaling during high light stress using microarray analyses. They found that H 2 O 2 was not only heavily involved in signaling in high light stress, but also salinity, water deficit, heat and cold stress. H 2 O 2 was a key regulator of small and 70 kD heat shock proteins and many genes of the anthocyanin metabolic pathway. Anthocyanins appear to play an important role as antioxidants in plants. A specific UDP-glycosyltransferase (UGT74E2) was highly regulated by H 2 O 2 . In a subsequent study [ 93 ], UGT74E2 responded quickly to H 2 O 2 and glycosylated indole-3-butyric acid (IBA) modifying auxin homeostasis, plant morphology and improving stress tolerance to salinity and water deficit. Furthermore, auxin was found to interact with ABA, increasing the ABA sensitivity of the plant. Silencing a poly(ADP-ribose) polymerase improved high light stress tolerance in Arabidopsis [ 94 , 95 ]. Part of the improved abiotic stress tolerance was ascribed to improved energy-use efficiency and reduced oxidative stress [ 94 , 95 ].
Kusano et al. [ 96 ] conducted a time-series experiment on the effects of UV-B light on Arabidopsis using both metabolomics and transcriptomics analyses. They found that plants responded in two phases with an upregulation of primary metabolites in the first phase and the induction of protective secondary metabolites, especially phenolics, in the second phase. The induction of phenolics corresponded to transcripts involved in the phenylpropanoid pathway, but the transcripts for primary metabolism were less consistent indicating that this pathway may be regulated by other mechanisms (e.g. kinases).
The transcriptomic response to drought can vary with the time of day [ 97 ]. These responses seem to interact with hormonal and other stress pathways that naturally vary during the course of the day. A smaller set of core genes were identified that responded at all times of the day. This set was compared to two previous studies and was whittled down to just 19 genes, including a NF-YB transcription factor, several PP2Cs, a CIPK7, and a sulfate transporter.
Drought stress studies and microarray analyses of three different genotypes of poplar clones grown in two different locations revealed epigenetic regulation to the environment [ 98 ]. The tree clones that had a longer history in the environment showed greater changes in DNA methylation, thereby influencing their response to drought.
Shoot tip growth of grapevines was found to be much more sensitive to osmotic stress than gene expression in a time-series experiment of the effects of gradual osmotic stress on grapevine [ 27 ]. Proteomics data indicated that changes in protein expression preceded and were not well correlated with gene expression (G.R. Cramer, unpublished results). The integration of transcriptomics data and metabolomics data indicated distinct differences of the responses of salinity and an isosmotic water deficit [ 27 ]. Drought-stressed plants induced greater responses in processes needed for osmotic adjustment and protection against ROS and photoinhibition. Salinity induced greater responses in processes involved in energy metabolism, ion transport, protein synthesis and protein fate. A comparison to similar short-term stresses [ 11 ] indicated that a gradual, chronic stress response was more complex than an acute stress response.
The effect of water-deficit on Cabernet Sauvignon berries (a red wine grape) in the field was studied using transcriptomics, proteomics and metabolomics [ 99 – 102 ]. Integrated analyses confirmed that the phenylpropanoid pathway (including anthocyanin and stilbene biosynthesis) was upregulated by water deficit in a tissue-specific manner in the skins of the berries. Other metabolic pathways in the berries were affected by water deficit including ABA, amino acid, carotenoid, lipid, sugar and acid metabolism. Most of these changes were associated with improved quality characteristics of the fruit.
Likewise, Zamboni et al. [ 103 ] investigated berry development and withering in grapevine at the transcriptomics, proteomics and metabolomics levels. A multistep hypothesis-free approach from four developmental stages and three withering intervals, with integration achieved using a hierarchical clustering strategy (multivariate O2PLS technique), identified stage-specific functional networks of linked transcripts, proteins and metabolites, providing important insights into the key molecular processes that determine wine quality. A hypothesis-driven approach identified transcript, protein and metabolite variables involved in the molecular events underpinning withering, which predominantly reflected a general stress response. Berry ripening and withering are characterized by the accumulation of secondary metabolites such as acylated anthocyanins, but withering also involves the activation of osmotic and oxidative stress response genes and the production of stilbenes and taxifolin.
Usadel et al. [ 104 ] investigated the effects of cold temperatures over time using transcriptomics, metabolomics and enzyme activities. They found some enzyme activities and metabolites changed rapidly, whereas others changed more slowly. The early changes (6 h) in enzyme activities were poorly correlated with transcript abundance, but after 78 h these correlations were greatly improved. Much of the long-term changes in metabolism could be ascribed to the CBF regulon.
Caldana et al. [ 105 ] conducted a complex time-series experiment (22 time points) with differing temperatures and light intensities using both metabolomics and transcriptomics analyses. This high-resolution time series experiment revealed that metabolic activities respond more quickly than transcriptional activities, indicating a disconnect between metabolism and transcription in the early phases of stress response and indicating that enzymatic activities may play a significant role. There were common metabolic responses to the changing environment within 1 h of the change including a decrease in energy metabolism and translation and an increase in the transcription of genes involved in signaling cascades. At later time points, condition-dependent metabolism was revealed. For example, protein degradation and energy metabolism derived from amino acids occurred in warm temperatures and darkness. Amino acid catabolism appears to fuel the TCA cycle in the absence of photosynthesis.
Yun et al. [ 106 ] characterized the response of rice to a mild chilling stress (10°C). They found that transcriptional regulation consisted of three dynamic and complex phases over 96 h. The early transcriptional phase appeared to be triggered by oxidative signals (H 2 O 2 ) and lead to the subsequent induction of cellular defense and rescue mechanisms. Combining temporal co-expression data from microarrays with promoter motif enrichment analyses and oxidative responses, transcriptional regulatory network models for the different response phases were constructed. A bZIP-TGA transcription factor module (as1/ocs/TGA), one of seven transcription factor modules, was the most connected regulatory module in phase one. Each of the transcription factor modules consisted of clusters of transcription factors exhibiting combinatorial control of the chilling regulon. The speed of the response of this network was associated with chilling tolerance. Chilling-resistant genotypes had a much more rapid and pronounced response of this transcriptional regulatory network than chilling-sensitive genotypes. In addition, the transcription factors identified in this study were located within known growth and stress QTLs in the rice genome.
Integration of omics analysis identifies molecular networks functioning in abiotic stress responses
Integrated omics analyses have markedly increased our understanding of plant responses to various stresses. These analyses are important for comprehensive analyses of abiotic stress responses, especially the final steps of stress signal transduction pathways.
Integrated analyses of the transcriptome and the metabolome successfully demonstrate connections between genes and metabolites, elucidating a wide range of signal output from ABA under dehydration [ 107 ] and the DREB1/CBF transcription factors in response to low temperature [ 108 , 109 ]. Metabolite profiling reveals that ABA accumulates during dehydration, regulating the accumulation of various amino acids and sugars such as glucose and fructose. In particular, the dehydration-inducible accumulation of BCAAs (branch-chain amino acids), saccharopine, proline, and agmatine are correlated with the dehydration-inducible expression of their key biosynthetic genes ( BCAT2 , LKR/SDH , P5CS1 , and ADC2 , respectively), which are regulated by endogenous ABA [ 107 ]. In addition, metabolome analysis of transgenic Arabidopsis overexpressing DREB1A/CBF3 reveals that there is a striking similarity between the low-temperature regulated metabolome (monosaccharides, disaccharides, oligosaccharides and sugar alcohols) and that regulated by the DREB1A/CBF3 transcription factor [ 108 , 109 ]. In particular, the low-temperature-inducible accumulation of galactinol and raffinose is correlated with the expression of the Gols3 gene, which is a direct target of DREB1A/CBF3 [ 108 , 109 ]. Maruyama et al. [ 109 ] also analyzed DREB2A overexpression, which did not increase the level of any low-temperature regulated metabolites in transgenic plants. Overexpression of DREB2A-CA in transgenic plants increased their tolerance to dehydration stress, but only slightly increased their tolerance to freezing stress [ 50 ]. These results indicate that the increased tolerance to freezing stress in transgenic plants overexpressing DREB1A may depend on the accumulation of low-temperature regulated metabolites, especially sucrose, raffinose, galactinol, and myo -inositol. Similarly, transcriptomics and metabolomics analyses of PSEUDO RESPONSE REGULATOR (PRR) arrhythmic triple mutant revealed that the DREB1A/CBF gene and raffinose amounts appear to be regulated by the circadian clock, varying between day and night as if in anticipation of the colder night temperatures [ 110 ].
Comparing metabolomics between dehydration, salinity, light, heat or low temperature stress have identified metabolites that are generally important in abiotic stress responses or are specific to each stress [ 27 , 95 , 105 , 111 , 112 ]. In a metabolite profiling study of Arabidopsis responses to combined dehydration and heat stresses [ 95 ], heat stress reduced the toxicity of proline, indicating that during the more severe combined stress treatment, sucrose replaces proline in plants as the major osmoprotectant. Comparative metabolite analysis between Arabidopsis responding to heat shock and cold shock revealed that the majority of metabolites in response to heat shock overlapped with those produced in response to cold shock [ 109 , 113 ]. These results indicate that a metabolic network of compatible solutes includes proline, monosaccharides (glucose and fructose), galactinol, and raffinose, which have an important role in tolerance to temperature stress. Wienkoop et al. [ 112 ] identified a RNA-binding protein (ATGRP7) that increased in response to low temperature stress and decreased in response to high temperature stress. Its abundance was significantly correlated with glutamine and proline concentrations. While raffinose and galactinol concentrations were significant markers for temperature responses, their response was independent of the responses of ATGRP7, proline and glutamine.
Transcriptomics, metabolomics and enzyme activities were integrated in a comprehensive study of K deficiency [ 114 ]. Carbon and nitrogen metabolism were significantly affected by K deficiency. This integrated approach pinpointed that pyruvate kinase activity (not transcription) was inhibited directly by K deficiency and was primarily responsible for the metabolic disorders observed.
Systematic application of omics technologies has contributed to the development of stress-tolerant crops in the field
Many genes affect stress tolerance, but few of the identified genes have proven useful in the field. Due to the complexity of stress interactions and stress responses, relevant phenotyping needs to be performed (including field experiments) in abiotic stress studies if we are to make significant progress [ 113 ]. The following studies are discussed to highlight good examples of systems biology and omics approaches that have been used to identify key genes regulating stress tolerance and then followed with validation of those responses and phenotypes in multiple experiments including field conditions.
A SNAC1 gene was identified from microarray experiments of stress treatments on rice [ 115 ]. SNAC1 is a NAC transcription factor that induces the expression of a number of stress-tolerance genes and improves the drought and salt tolerance of rice in the field. The transgenic plants exhibited increased sensitivity to ABA and reduced water loss. In another drought stress study, a LEA (late embryogenesis abundant) gene was identified from microarray experiments of rice and was transformed and tested in the field under drought conditions through the T3 generation [ 116 ]. Spikelet fertility appears to be the main factor contributing to improved yields under drought conditions.
An exhaustive screen of greater than 1500 transcription factors in Arabidopsis identified approximately 40 transcription factors that when overexpressed, improved stress tolerance [ 117 ]. One of these transcription factors NF-YB1 was further characterized and shown to display significant drought tolerance in Arabidopsis. Microarray data of this overexpressing line showed few differences in gene expression and the genes identified were not known previously to be involved in drought tolerance. This functional genomics approach provided a new strategy for improving drought tolerance in plants. A homolog of NF-YB1 was cloned in maize (ZmNF-YB2), overexpressed and tested for drought tolerance in the greenhouse and field plots. The transgenic maize lines were more drought tolerant having increased chlorophyll content, photosynthesis, stomatal conductance and grain yields. One line consistently had more than 50% yield improvement in drought conditions over two different years.
Oh et al. [ 118 ] used microarrays to identify 42 AP2 transcription factors whose expressions were affected by stress. Two of these transcription factors, AP37 and AP59 were functionally characterized. The two transcription factors are closely related but have distinct differences in affecting rice phenotype. AP37 responded to drought, salinity, cold and ABA; over-expression improved stress tolerance to all three environmental conditions. AP59 responded to drought and salinity, but not cold or ABA, and improved stress tolerance to drought and salinity only. Both overexpressing lines showed improved photosynthetic efficiency under stress conditions. Overexpression of the transcription factors induced common and distinct sets of genes. T5 homozygous overexpressing lines of AP59, but not AP37, had yield penalties under normal paddy conditions in the field, whereas AP37 overexpressing lines, but not AP59, had enhanced yields under drought conditions in the field. The reduced yields of the overexpressing lines of AP59 were attributed to effects on spikelet development. This study emphasizes the point that it is important to characterize gene effects on yield under field conditions.
Mapping stress responses has provided new insights and identified gaps in our knowledge of abiotic stress responses
From a meta-analysis of drought-stress related papers from the last 15 years, a complex model for plant responses to drought stress was produced [ 12 ]. This model details the interactions of sugars, ROS/RNS, hormones (ABA, ethylene, auxins, cytokinins, salicylic acid, gibberellin and brassinosteroids) and nitrogen metabolism. It highlights the highly complex nature of stress responses.
From this review, we have constructed a simplified working model summarizing some of the known plant signaling responses to abiotic stress (Figure 2 ). Much of the signaling involves phosphorylation cascades that react quickly in the plant cell, emphasizing the need for proteomics data as well as transcriptomics data in future models. The PYR/PYL/RCAR-PP2C-SnRK2 pathway illustrates that protein phosphorylation and dephosphorylation are the most important factors in ABA signaling. Similar phosphorylation and dephosphorylation processes are involved in ethylene and other abiotic stress signaling pathways (Figure 2 ). Not all connections could be drawn in this two-dimensional figure without obscuring many other connections. For example, the interactions of ROS with abiotic stresses and hormones [ 32 ] are too complex to display here. In addition, the actual signaling response will be dependent upon the signaling pathway present in that organ, tissue or cell at the time of the response. One needs to use more sophisticated bioinformatics programs like Cytoscape [ 119 ] and its plug-ins to visualize the interactions comprehensively in two dimensional or three-dimensional space [ 120 ] or with time series views [ 121 ], which would allow these data to be viewed in four dimensions.
Although there are still some technological issues that must be solved to produce a complete picture of protein phosphorylation, several technologies have been developed for the large-scale analysis of phosphoproteins, known as 'phosphoproteomics' [ 122 ]. Mass spectrometry analyses have identified thousands of phosphoproteins in Arabidopsis, rice, and Medicago truncatula [ 123 – 125 ]. In addition, two studies have reported ABA-responsive changes in the phosphoproteome [ 126 , 127 ]. Phosphoproteomics analyses of mutants for abiotic stress signaling (e.g. PP2C or SnRK) will identify the relevant network of protein phosphorylation events in abiotic stress signaling.
Transcriptome analysis technologies have advanced to the point where high-through-put DNA sequencers and high-density microarrays such as tiling arrays are readily available. These technologies provide new opportunities to analyze noncoding RNAs and can clarify aspects of epigenetic regulation of gene expression [ 128 , 129 ]. Similar approaches [ 130 , 131 ] have revealed the global transcriptomes of plants exposed to abiotic stresses such as dehydration, cold, heat, high-salinity, osmotic stress, and ABA. These analyses indicate that these stresses increase or decrease transcript abundance from not only previously identified stress-responsive genes, but also from thousands of unannotated non-protein-coding regions. Matsui et al. [ 130 ] estimated that approximately 80% of previously unannotated upregulated transcripts arise from antisense strands of sense transcripts. There was a significant linear correlation between the expression ratios (stress-treated/untreated) of the sense transcripts and the ratios of the antisense transcripts. Interestingly, the data suggested that such stress-responsive antisense transcripts are derived from antisense strands of the stress-responsive genes, RD29A and CYP707A1 . Clearly, transcriptional regulation is far more complicated than we previously imagined. Whether or not such antisense transcripts have biological functions is an important issue that remains to be resolved.
Much more research is required in order to fully map plant responses to abiotic stress. The nature of the pathway responses will vary and is highly dependent on the species, organ, tissue, cell type, developmental stage of the plant, the stress or stresses affecting the plant, the level and duration of the stress. Despite the vast amount of research collected on abiotic stress in the last decade, there are still significant gaps in our knowledge. We still do not understand completely how plants perceive stress. We don't know all of the receptors and their sites of action (organs, tissues and cellular components). While we know a lot about downstream signaling (i.e. transcriptional pathways), we know very little about the primary signaling (i.e. proteomics). Most of the literature on abiotic stress responses in plants is based upon transcriptomics data rather than proteomics data. This is not surprising as transcriptomics technology is more advanced, easier to perform and less expensive. However, transcriptomics analyses are insufficient as there is an overall poor correlation of transcriptomics profiles with proteomics profiles [ 101 , 132 , 133 ] or enzyme activities [ 104 , 114 ]. There are only a few studies describing phosphorylation cascades and other post-translation modification activities in response to stress [ 134 ]. Recent efforts to map the hormone [ 126 , 127 ] and light-regulated [ 135 ] phosphorylomes are good first steps. Finally, we need better tools to facilitate systems biology analyses especially in the area of bioinformatics. Transcriptomics data can be collected in a matter of days or weeks, but the data analyses often take more than a year.
Conclusions
We have made great progress in understanding the responses of plants to abiotic stress. There are inherent physical, morphological and molecular limitations to the plant's ability to respond to stress. Systems biology approaches have given us a more holistic view of the molecular responses. Transcriptomics studies are well advanced, but proteomics analyses are lagging behind, especially the study of post-translational modifications. Plant responses to abiotic stress are dynamic and complex. The integration of multiple omics studies has revealed new areas of interactions and regulation. Time series experiments have revealed the kinetics of stress responses, identifying multiple response phases involving core sets of genes and condition-dependent changes. One consistent trend in response to abiotic stress is the early down regulation of energy metabolism and protein synthesis. This may indicate a conservation of energy by the plant and may reflect a shift from plant growth to protective mechanisms. In many examples presented in this review, ABA signaling mediates the plant responses to abiotic stress. Co-expression analyses are useful in that they have revealed key regulatory hubs that can be manipulated to produce different phenotypes. To get a comprehensive understanding of plant responses to abiotic stress, more extensive mapping of these responses at the organ, tissue and cellular level are needed. Such network analyses need to be extended to the proteomics and enzyme activities levels. Models need to be constructed and linked to phenotypic traits. The linkage of key regulatory hubs to phenotypic traits will allow for more rapid progress in the genetic manipulation and production of crop plants. Current progress is exemplified by the identification and validation of several key genes that improved stress tolerance of crops in the field. It is expected that progress in the plant sciences and systems biology will continue to accelerate in the near future.
Kitano H: Systems biology: a brief overview. Science. 2002, 295 (5560): 1662-1664. 10.1126/science.1069492.
PubMed CAS Google Scholar
Feist AM, Palsson BO: The growing scope of applications of genome-scale metabolic reconstructions using Escherichia coli. Nat Biotechnol. 2008, 26 (6): 659-667. 10.1038/nbt1401.
PubMed CAS PubMed Central Google Scholar
Bemis SM, Torii KU: Autonomy of cell proliferation and developmental programs during Arabidopsis aboveground organ morphogenesis. Dev Biol. 2007, 304 (1): 367-381. 10.1016/j.ydbio.2006.12.049.
Riefler M, Novak O, Strnad M, Schmulling T: Arabidopsis cytokinin receptor mutants reveal functions in shoot growth, leaf senescence, seed size, germination, root development, and cytokinin metabolism. Plant Cell. 2006, 18 (1): 40-54. 10.1105/tpc.105.037796.
Gibeaut DM, Hulett J, Cramer GR, Seemann JR: Maximal biomass of Arabidopsis thaliana using a simple, low-maintenance hydroponic method and favorable environmental conditions. Plant Physiol. 1997, 115: 317-319. 10.1104/pp.115.2.317.
Boyer JS: Plant productivity and environment. Science. 1982, 218 (4571): 443-448. 10.1126/science.218.4571.443.
Lobell DB, Schlenker W, Costa-Roberts J: Climate trends and global crop production since 1980. Science. 2011, 333 (6042): 616-620. 10.1126/science.1204531.
Skirycz A, Inze D: More from less: plant growth under limited water. Curr Opin Biotechnol. 2010, 21 (2): 197-203. 10.1016/j.copbio.2010.03.002.
Cramer GR: Abiotic stress & plant responses from the whole vine to the genes. Aust J Grape Wine Res. 2010, 16: 86-93.
CAS Google Scholar
Dinneny JR, Long TA, Wang JY, Jung JW, Mace D, Pointer S, Barron C, Brady SM, Schiefelbein J, Benfey PN: Cell identity mediates the response of Arabidopsis roots to abiotic stress. Science. 2008, 320 (5878): 942-945. 10.1126/science.1153795.
Tattersall EA, Grimplet J, Deluc L, Wheatley MD, Vincent D, Osborne C, Ergul A, Lomen E, Blank RR, Schlauch KA, Cushman JC, Cramer GR: Transcript abundance profiles reveal larger and more complex responses of grapevine to chilling compared to osmotic and salinity stress. Funct Integr Genomics. 2007, 7 (4): 317-333. 10.1007/s10142-007-0051-x.
Pinheiro C, Chaves MM: Photosynthesis and drought: can we make metabolic connections from available data?. J Exp Bot. 2011, 62 (3): 869-882. 10.1093/jxb/erq340.
Boyer JS: Evans Review: Cell wall biosynthesis and the molecular mechanism of plant enlargement. Funct Plant Biol. 2009, 36 (5): 383-394. 10.1071/FP09048.
Parent B, Hachez C, Redondo E, Simonneau T, Chaumont F, Tardieu F: Drought and abscisic acid effects on aquaporin content translate into changes in hydraulic conductivity and leaf growth rate: a trans-scale approach. Plant Physiol. 2009, 149 (4): 2000-2012. 10.1104/pp.108.130682.
Boursiac Y, Boudet J, Postaire O, Luu DT, Tournaire-Roux C, Maurel C: Stimulus-induced downregulation of root water transport involves reactive oxygen species-activated cell signalling and plasma membrane intrinsic protein internalization. Plant J. 2008, 56 (2): 207-218. 10.1111/j.1365-313X.2008.03594.x.
Vandeleur RK, Mayo G, Shelden MC, Gilliham M, Kaiser BN, Tyerman SD: The role of PIP aquaporins in water transport through roots: diurnal and drought stress responses reveal different strategies between isohydric and anisohydric cultivars of grapevine. Plant Physiol. 2008
Google Scholar
Nardini A, Lo GMA, Salleo S: Refilling embolized xylem conduits: is it a matter of phloem unloading?. Plant Sci. 2011, 180 (4): 604-611. 10.1016/j.plantsci.2010.12.011.
Hummel I, Pantin F, Sulpice R, Piques M, Rolland G, Dauzat M, Christophe A, Pervent M, Bouteille M, Stitt M, Gibon Y, Muller B: Arabidopsis plants acclimate to water deficit at low cost through changes of carbon usage: an integrated perspective using growth, metabolite, enzyme, and gene expression analysis. Plant Physiol. 2010, 154 (1): 357-372. 10.1104/pp.110.157008.
Cramer GR, Alberico GJ, Schmidt C: Leaf expansion limits dry matter accumulation of salt-stressed maize. Aust J Plant Physiol. 1994, 21: 663-674. 10.1071/PP9940663.
Nonami H, Wu YJ, Boyer JS: Decreased growth-induced water potential: primary cause of growth inhibition at low water potentials. Plant Physiol. 1997, 114 (2): 501-509.
Tang AC, Boyer JS: Growth-induced water potentials and the growth of maize leaves. J Exp Bot. 2002, 53 (368): 489-503. 10.1093/jexbot/53.368.489.
Good AG, Zaplachinski ST: The effects of drought stress on free amino acid accumulation and protein synthesis in brassica napus. Physiol Plant. 1994, 90 (1): 9-14. 10.1111/j.1399-3054.1994.tb02185.x.
Vincent D, Ergul A, Bohlman MC, Tattersall EA, Tillett RL, Wheatley MD, Woolsey R, Quilici DR, Joets J, Schlauch K, Schooley DA, Cushman JC, Cramer GR: Proteomic analysis reveals differences between Vitis vinifera L. cv. Chardonnay and cv. Cabernet Sauvignon and their responses to water deficit and salinity. J Exp Bot. 2007, 58 (7): 1873-1892. 10.1093/jxb/erm012.
Ben-Zioni A, Itai C, Vaadia Y: Water and salt stresses, kinetin and protein synthesis in tobacco leaves. Plant Physiol. 1967, 42: 361-365. 10.1104/pp.42.3.361.
Dhindsa RS, Cleland RE: Water stress and protein synthesis: I. Differential inhibition of protein synthesis. Plant Physiol. 1975, 55 (4): 778-781. 10.1104/pp.55.4.778.
Liu JX, Howell SH: Endoplasmic reticulum protein quality control and its relationship to environmental stress responses in plants. Plant Cell. 2010, 22 (9): 2930-2942. 10.1105/tpc.110.078154.
Cramer GR, Ergul A, Grimplet J, Tillett RL, Tattersall EA, Bohlman MC, Vincent D, Sonderegger J, Evans J, Osborne C, Quilici D, Schlauch KA, Schooley DA, Cushman JC: Water and salinity stress in grapevines: early and late changes in transcript and metabolite profiles. Funct Integr Genomics. 2007, 7 (2): 111-134. 10.1007/s10142-006-0039-y.
Kilian J, Whitehead D, Horak J, Wanke D, Weinl S, Batistic O, D'Angelo C, Bornberg-Bauer E, Kudla J, Harter K: The AtGenExpress global stress expression data set: protocols, evaluation and model data analysis of UV-B light, drought and cold stress responses. Plant J. 2007, 50 (2): 347-363. 10.1111/j.1365-313X.2007.03052.x.
Takahashi S, Seki M, Ishida J, Satou M, Sakurai T, Narusaka M, Kamiya A, Nakajima M, Enju A, Akiyama K, Yamaguchi-Shinozaki K, Shinozaki K: Monitoring the expression profiles of genes induced by hyperosmotic, high salinity, and oxidative stress and abscisic acid treatment in Arabidopsis cell culture using a full-length cDNA microarray. Plant Mol Biol. 2004, 56 (1): 29-55. 10.1007/s11103-004-2200-0.
Molassiotis A, Fotopoulos V: Oxidative and nitrosative signaling in plants: two branches in the same tree?. Plant Signal Behav. 2011, 6 (2): 210-214. 10.4161/psb.6.2.14878.
Wilkinson S, Davies WJ: Drought, ozone, ABA and ethylene: new insights from cell to plant to community. Plant Cell Environ. 2009
Mittler R, Vanderauwera S, Suzuki N, Miller G, Tognetti VB, Vandepoele K, Gollery M, Shulaev V, Van BF: ROS signaling: the new wave?. Trends Plant Sci. 2011, 16 (6): 300-309. 10.1016/j.tplants.2011.03.007.
Goda H, Sasaki E, Akiyama K, Maruyama-Nakashita A, Nakabayashi K, Li W, Ogawa M, Yamauchi Y, Preston J, Aoki K, Kiba T, Takatsuto S, Fujioka S, Asami T, Nakano T, Kato H, Mizuno T, Sakakibara H, Yamaguchi S, Nambara E, Kamiya Y, Takahashi H, Hirai MY, Sakurai T, Shinozaki K, Saito K, Yoshida S, Shimada Y: The AtGenExpress hormone and chemical treatment data set: experimental design, data evaluation, model data analysis and data access. Plant J. 2008, 55 (3): 526-542. 10.1111/j.1365-313X.2008.03510.x.
Hubbard KE, Nishimura N, Hitomi K, Getzoff ED, Schroeder JI: Early abscisic acid signal transduction mechanisms: newly discovered components and newly emerging questions. Genes Dev. 2010, 24 (16): 1695-1708. 10.1101/gad.1953910.
Kim TH, Bohmer M, Hu H, Nishimura N, Schroeder JI: Guard cell signal transduction network: advances in understanding abscisic acid, CO2, and Ca2+ signaling. Annu Rev Plant Biol. 2010, 61: 561-591. 10.1146/annurev-arplant-042809-112226.
Chinnusamy V, Gong Z, Zhu JK: Abscisic acid-mediated epigenetic processes in plant development and stress responses. J Integr Plant Biol. 2008, 50 (10): 1187-1195. 10.1111/j.1744-7909.2008.00727.x.
Umezawa T: Systems biology approaches to abscisic acid signaling. J Plant Res. 2011, 124 (4): 539-548. 10.1007/s10265-011-0418-x.
Ma Y, Szostkiewicz I, Korte A, Moes D, Yang Y, Christmann A, Grill E: Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science. 2009, 324 (5930): 1064-1068.
Park SY, Fung P, Nishimura N, Jensen DR, Fujii H, Zhao Y, Lumba S, Santiago J, Rodrigues A, Chow TF, Alfred SE, Bonetta D, Finkelstein R, Provart NJ, Desveaux D, Rodriguez PL, McCourt P, Zhu JK, Schroeder JI, Volkman BF, Cutler SR: Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science. 2009, 324 (5930): 1068-1071.
Leung J, Giraudat J: Abscisic acid signal transduction. Annu Rev Plant Physiol Plant Mol Biol. 1998, 49: 199-222. 10.1146/annurev.arplant.49.1.199.
Yoshida R, Hobo T, Ichimura K, Mizoguchi T, Takahashi F, Aronso J, Ecker JR, Shinozaki K: ABA-activated SnRK2 protein kinase is required for dehydration stress signaling in Arabidopsis. Plant Cell Physiol. 2002, 43 (12): 1473-1483. 10.1093/pcp/pcf188.
Mustilli AC, Merlot S, Vavasseur A, Fenzi F, Giraudat J: Arabidopsis OST1 protein kinase mediates the regulation of stomatal aperture by abscisic acid and acts upstream of reactive oxygen species production. Plant Cell. 2002, 14 (12): 3089-3099. 10.1105/tpc.007906.
Umezawa T, Nakashima K, Miyakawa T, Kuromori T, Tanokura M, Shinozaki K, Yamaguchi-Shinozaki K: Molecular basis of the core regulatory network in aba responses: sensing, signaling and transport. Plant Cell Physiol. 2010, 51 (11): 1821-1839. 10.1093/pcp/pcq156.
Yamaguchi-Shinozaki K, Shinozaki K: Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu Rev Plant Biol. 2006, 57: 781-803. 10.1146/annurev.arplant.57.032905.105444.
Fujita Y, Fujita M, Satoh R, Maruyama K, Parvez MM, Seki M, Hiratsu K, Ohme-Takagi M, Shinozaki K, Yamaguchi-Shinozaki K: AREB1 is a transcription activator of novel ABRE-dependent ABA signaling that enhances drought stress tolerance in Arabidopsis. Plant Cell. 2005, 17 (12): 3470-3488. 10.1105/tpc.105.035659.
Kang JY, Choi HI, Im MY, Kim SY: Arabidopsis basic leucine zipper proteins that mediate stress-responsive abscisic Acid signaling. Plant Cell. 2002, 14 (2): 343-357. 10.1105/tpc.010362.
Yoshida T, Fujita Y, Sayama H, Maruyama K, Mizoi J, Shinozaki K, Yamaguchi-Shinozaki K: AREB1, AREB2, and ABF3 are master transcription factors that cooperatively regulate ABRE-dependent ABA signaling involved in drought stress tolerance and require ABA for full activation. Plant J. 2010, 61 (4): 672-685. 10.1111/j.1365-313X.2009.04092.x.
Fowler S, Thomashow MF: Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway. Plant Cell. 2002, 14 (8): 1675-1690. 10.1105/tpc.003483.
Maruyama K, Sakuma Y, Kasuga M, Ito Y, Seki M, Goda H, Shimada Y, Yoshida S, Shinozaki K, Yamaguchi-Shinozaki K: Identification of cold-inducible downstream genes of the Arabidopsis DREB1A/CBF3 transcriptional factor using two microarray systems. Plant J. 2004, 38 (6): 982-993. 10.1111/j.1365-313X.2004.02100.x.
Sakuma Y, Maruyama K, Osakabe Y, Qin F, Seki M, Shinozaki K, Yamaguchi-Shinozaki K: Functional analysis of an Arabidopsis transcription factor, DREB2A, involved in drought-responsive gene expression. Plant Cell. 2006, 18 (5): 1292-1309. 10.1105/tpc.105.035881.
Stepanova AN, Alonso JM: Ethylene signaling and response: where different regulatory modules meet. Curr Opin Plant Biol. 2009, 12 (5): 548-555. 10.1016/j.pbi.2009.07.009.
Yoo SD, Cho Y, Sheen J: Emerging connections in the ethylene signaling network. Trends Plant Sci. 2009, 14 (5): 270-279. 10.1016/j.tplants.2009.02.007.
Morgan PW, Drew MC: Ethylene and plant responses to stress. Physiol Plant. 1997, 100 (3): 620-630. 10.1111/j.1399-3054.1997.tb03068.x.
Zhang M, Leng P, Zhang G, Li X: Cloning and functional analysis of 9-cis-epoxycarotenoid dioxygenase (NCED) genes encoding a key enzyme during abscisic acid biosynthesis from peach and grape fruits. J Plant Physiol. 2009, 166 (12): 1241-1252. 10.1016/j.jplph.2009.01.013.
Sun L, Zhang M, Ren J, Qi J, Zhang G, Leng P: Reciprocity between abscisic acid and ethylene at the onset of berry ripening and after harvest. BMC Plant Biol. 2010, 10: 257-10.1186/1471-2229-10-257.
PubMed PubMed Central Google Scholar
Ophir R, Pang X, Halaly T, Venkateswari J, Lavee S, Galbraith D, Or E: Gene-expression profiling of grape bud response to two alternative dormancy-release stimuli expose possible links between impaired mitochondrial activity, hypoxia, ethylene-ABA interplay and cell enlargement. Plant Mol Biol. 2009, 71 (4-5): 403-423. 10.1007/s11103-009-9531-9.
Petranovic D, Tyo K, Vemuri GN, Nielsen J: Prospects of yeast systems biology for human health: integrating lipid, protein and energy metabolism. FEMS Yeast Res. 2010, 10 (8): 1046-1059. 10.1111/j.1567-1364.2010.00689.x.
Zaborske JM, Wu X, Wek RC, Pan T: Selective control of amino acid metabolism by the GCN2 eIF2 kinase pathway in Saccharomyces cerevisiae. BMC Biochem. 2010, 11: 29-10.1186/1471-2091-11-29.
Staschke KA, Dey S, Zaborske JM, Palam LR, McClintick JN, Pan T, Edenberg HJ, Wek RC: Integration of general amino acid control and target of rapamycin (TOR) regulatory pathways in nitrogen assimilation in yeast. J Biol Chem. 2010, 285 (22): 16893-16911. 10.1074/jbc.M110.121947.
Smeekens S, Ma J, Hanson J, Rolland F: Sugar signals and molecular networks controlling plant growth. Curr Opin Plant Biol. 2010, 13 (3): 274-279.
Hey SJ, Byrne E, Halford NG: The interface between metabolic and stress signalling. Ann Bot. 2010, 105 (2): 197-203. 10.1093/aob/mcp285.
Deprost D, Yao L, Sormani R, Moreau M, Leterreux G, Nicolai M, Bedu M, Robaglia C, Meyer C: The Arabidopsis TOR kinase links plant growth, yield, stress resistance and mRNA translation. EMBO Rep. 2007, 8 (9): 864-870. 10.1038/sj.embor.7401043.
Lageix S, Lanet E, Pouch-Pelissier MN, Espagnol MC, Robaglia C, Deragon JM, Pelissier T: Arabidopsis eIF2alpha kinase GCN2 is essential for growth in stress conditions and is activated by wounding. BMC Plant Biol. 2008, 8: 134-10.1186/1471-2229-8-134.
Baena-Gonzalez E, Rolland F, Thevelein JM, Sheen J: A central integrator of transcription networks in plant stress and energy signalling. Nature. 2007, 448 (7156): 938-942. 10.1038/nature06069.
Baena-Gonzalez E, Sheen J: Convergent energy and stress signaling. Trends Plant Sci. 2008, 13 (9): 474-482. 10.1016/j.tplants.2008.06.006.
Jossier M, Bouly JP, Meimoun P, Arjmand A, Lessard P, Hawley S, Grahame HD, Thomas M: SnRK1 (SNF1-related kinase 1) has a central role in sugar and ABA signalling in Arabidopsis thaliana. Plant J. 2009, 59 (2): 316-328. 10.1111/j.1365-313X.2009.03871.x.
Obertello M, Krouk G, Katari MS, Runko SJ, Coruzzi GM: Modeling the global effect of the basic-leucine zipper transcription factor 1 (bZIP1) on nitrogen and light regulation in Arabidopsis. BMC Systems Biol. 2010, 4: 111-10.1186/1752-0509-4-111.
Kang SG, Price J, Lin PC, Hong JC, Jang JC: The arabidopsis bZIP1 transcription factor is involved in sugar signaling, protein networking, and DNA binding. Mol Plant. 2010, 3 (2): 361-373. 10.1093/mp/ssp115.
Osuna D, Usadel B, Morcuende R, Gibon Y, Blasing OE, Hohne M, Gunter M, Kamlage B, Trethewey R, Scheible WR, Stitt M: Temporal responses of transcripts, enzyme activities and metabolites after adding sucrose to carbon-deprived Arabidopsis seedlings. Plant J. 2007, 49 (3): 463-491. 10.1111/j.1365-313X.2006.02979.x.
Zhou L, Jang JC, Jones TL, Sheen J: Glucose and ethylene signal transduction crosstalk revealed by an Arabidopsis glucose-insensitive mutant. Proc Natl Acad Sci USA. 1998, 95 (17): 10294-10299. 10.1073/pnas.95.17.10294.
Arenas-Huertero F, Arroyo A, Zhou L, Sheen J, Leon P: Analysis of Arabidopsis glucose insensitive mutants, gin5 and gin6, reveals a central role of the plant hormone ABA in the regulation of plant vegetative development by sugar. Genes Dev. 2000, 14 (16): 2085-2096.
Finkelstein RR, Gibson SI: ABA and sugar interactions regulating development: cross-talk or voices in a crowd?. Curr Opin Plant Biol. 2002, 5 (1): 26-32. 10.1016/S1369-5266(01)00225-4.
Franco-Zorrilla JM, Martin AC, Leyva A, Paz-Ares J: Interaction between phosphate-starvation, sugar, and cytokinin signaling in Arabidopsis and the roles of cytokinin receptors CRE1/AHK4 and AHK3. Plant Physiol. 2005, 138 (2): 847-857. 10.1104/pp.105.060517.
Mita S, Suzuki-Fujii K, Nakamura K: Sugar-inducible expression of a gene for beta-amylase in Arabidopsis thaliana. Plant Physiol. 1995, 107 (3): 895-904. 10.1104/pp.107.3.895.
Thum KE, Shasha DE, Lejay LV, Coruzzi GM: Light- and carbon-signaling pathways. Modeling circuits of interactions. Plant Physiol. 2003, 132 (2): 440-452. 10.1104/pp.103.022780.
Cakir B, Agasse A, Gaillard C, Saumonneau A, Delrot S, Atanassova R: A grape ASR protein involved in sugar and abscisic acid signaling. Plant Cell. 2003, 15 (9): 2165-2180. 10.1105/tpc.013854.
Lecourieux F, Lecourieux D, Vignault C, Delrot S: A sugar inducible protein kinase, VvSK1, regulates hexose transport and sugar accumulation in grapevine cells. Plant Physiol. 2010, 52: 1096-1106.
Shen W, Reyes MI, Hanley-Bowdoin L: Arabidopsis protein kinases GRIK1 and GRIK2 specifically activate SnRK1 by phosphorylating its activation loop. Plant Physiol. 2009, 150 (2): 996-1005. 10.1104/pp.108.132787.
Tiessen A, Prescha K, Branscheid A, Palacios N, McKibbin R, Halford NG, Geigenberger P: Evidence that SNF1-related kinase and hexokinase are involved in separate sugar-signalling pathways modulating post-translational redox activation of ADP-glucose pyrophosphorylase in potato tubers. Plant J. 2003, 35 (4): 490-500. 10.1046/j.1365-313X.2003.01823.x.
Zheng Z, Xu X, Crosley RA, Greenwalt SA, Sun Y, Blakeslee B, Wang L, Ni W, Sopko MS, Yao C, Yau K, Burton S, Zhuang M, McCaskill DG, Gachotte D, Thompson M, Greene TW: The protein kinase SnRK2.6 mediates the regulation of sucrose metabolism and plant growth in Arabidopsis. Plant Physiol. 2010, 153 (1): 99-113. 10.1104/pp.109.150789.
Lenz T, Fischer JJ, Dreger M: Probing small molecule-protein interactions: A new perspective for functional proteomics. J Proteomics. 2011
Kaufmann K, Smaczniak C, de VS, Angenent GC, Karlova R: Proteomics insights into plant signaling and development. Proteomics. 2011, 11 (4): 744-755. 10.1002/pmic.201000418.
Usadel B, Obayashi T, Mutwil M, Giorgi FM, Bassel GW, Tanimoto M, Chow A, Steinhauser D, Persson S, Provart NJ: Co-expression tools for plant biology: opportunities for hypothesis generation and caveats. Plant Cell Environ. 2009, 32 (12): 1633-1651. 10.1111/j.1365-3040.2009.02040.x.
Hirai MY, Yano M, Goodenowe DB, Kanaya S, Kimura T, Awazuhara M, Arita M, Fujiwara T, Saito K: Integration of transcriptomics and metabolomics for understanding of global responses to nutritional stresses in Arabidopsis thaliana. Proc Natl Acad Sci USA. 2004, 101 (27): 10205-10210. 10.1073/pnas.0403218101.
Hirai MY, Sugiyama K, Sawada Y, Tohge T, Obayashi T, Suzuki A, Araki R, Sakurai N, Suzuki H, Aoki K, Goda H, Nishizawa OI, Shibata D, Saito K: Omics-based identification of Arabidopsis Myb transcription factors regulating aliphatic glucosinolate biosynthesis. Proc Natl Acad Sci USA. 2007, 104 (15): 6478-6483. 10.1073/pnas.0611629104.
Mao L, Van HJL, Dash S, Dickerson JA: Arabidopsis gene co-expression network and its functional modules. BMC Bioinformatics. 2009, 10: 346-10.1186/1471-2105-10-346.
Carrera J, Rodrigo G, Jaramillo A, Elena SF: Reverse-engineering the Arabidopsis thaliana transcriptional network under changing environmental conditions. Genome Biol. 2009, 10 (9): R96.-
Lorenz WW, Alba R, Yu YS, Bordeaux JM, Simoes M, Dean JF: Microarray analysis and scale-free gene networks identify candidate regulators in drought-stressed roots of loblolly pine (P. taeda L.). BMC Genomics. 2011, 12: 264-10.1186/1471-2164-12-264.
Weston DJ, Gunter LE, Rogers A, Wullschleger SD: Connecting genes, coexpression modules, and molecular signatures to environmental stress phenotypes in plants. BMC Systems Biol. 2008, 2: 16-10.1186/1752-0509-2-16.
Weston DJ, Karve AA, Gunter LE, Jawdy SS, Yang X, Allen SM, Wullschleger SD: Comparative physiology and transcriptional networks underlying the heat shock response in Populus trichocarpa, Arabidopsis thaliana and Glycine max. Plant Cell Environ. 2011
Sun X, Zou Y, Nikiforova V, Kurths J, Walther D: The complexity of gene expression dynamics revealed by permutation entropy. BMC Bioinformatics. 2010, 11: 607-10.1186/1471-2105-11-607.
Vanderauwera S, Zimmermann P, Rombauts S, Vandenabeele S, Langebartels C, Gruissem W, Inze D, Van BF: Genome-wide analysis of hydrogen peroxide-regulated gene expression in Arabidopsis reveals a high light-induced transcriptional cluster involved in anthocyanin biosynthesis. Plant Physiol. 2005, 139 (2): 806-821. 10.1104/pp.105.065896.
Tognetti VB, Van AO, Morreel K, Vandenbroucke K, van dCB, De CI, Chiwocha S, Fenske R, Prinsen E, Boerjan W, Genty B, Stubbs KA, Inze D, Van BF: Perturbation of indole-3-butyric acid homeostasis by the UDP-glucosyltransferase UGT74E2 modulates Arabidopsis architecture and water stress tolerance. Plant Cell. 2010, 22 (8): 2660-2679. 10.1105/tpc.109.071316.
Vanderauwera S, De BM, Van dSN, van dCB, Metzlaff M, Van BF: Silencing of poly(ADP-ribose) polymerase in plants alters abiotic stress signal transduction. Proc Natl Acad Sci USA. 2007, 104 (38): 15150-15155. 10.1073/pnas.0706668104.
De Block M, Verduyn C, De Brouwer D, Cornelissen M: Poly(ADP-ribose) polymerase in plants affects energy homeostasis, cell death and stress tolerance. Plant J. 2005, 41 (1): 95-106.
Kusano M, Tohge T, Fukushima A, Kobayashi M, Hayashi N, Otsuki H, Kondou Y, Goto H, Kawashima M, Matsuda F, Niida R, Matsui M, Saito K, Fernie AR: Metabolomics reveals comprehensive reprogramming involving two independent metabolic responses of Arabidopsis to UV-B light. Plant J. 2011, 67 (2): 354-369. 10.1111/j.1365-313X.2011.04599.x.
Wilkins O, Brautigam K, Campbell MM: Time of day shapes Arabidopsis drought transcriptomes. Plant J. 2010, 63 (5): 715-727. 10.1111/j.1365-313X.2010.04274.x.
Raj S, Brautigam K, Hamanishi ET, Wilkins O, Thomas BR, Schroeder W, Mansfield SD, Plant AL, Campbell MM: Clone history shapes Populus drought responses. Proc Natl Acad Sci USA. 2011
Grimplet J, Deluc LG, Tillett RL, Wheatley MD, Schlauch KA, Cramer GR, Cushman JC: Tissue-specific mRNA expression profiling in grape berry tissues. BMC Genomics. 2007, 8: 187-10.1186/1471-2164-8-187.
Deluc LG, Quilici DR, Decendit A, Grimplet J, Wheatley MD, Schlauch KA, Merillon JM, Cushman JC, Cramer GR: Water deficit alters differentially metabolic pathways affecting important flavor and quality traits in grape berries of Cabernet Sauvignon and Chardonnay. BMC Genomics. 2009, 10: 212-10.1186/1471-2164-10-212.
Grimplet J, Wheatley MD, Jouira HB, Deluc LG, Cramer GR, Cushman JC: Proteomic and selected metabolite analysis of grape berry tissues under well watered and water-deficit stress conditions. Proteomics. 2009, 9: 2503-2528. 10.1002/pmic.200800158.
Deluc LG, Decendit A, Papastamoulis Y, Merillon JM, Cushman JC, Cramer GR: Water Deficit Increases Stilbene Metabolism in Cabernet Sauvignon Berries. J Agric Food Chem. 2011, 59 (1): 289-297. 10.1021/jf1024888.
Zamboni A, Di Carli M, Guzzo F, Stocchero M, Zenoni S, Ferrarini A, Tononi P, Toffali K, Desiderio A, Lilley KS, Pe ME, Benvenuto E, Delledonne M, Pezzotti M: Identification of putative stage-specific grapevine berry biomarkers and omics data integration into networks. Plant Physiol. 2010, 154 (3): 1439-1459. 10.1104/pp.110.160275.
Usadel B, Blasing OE, Gibon Y, Poree F, Hohne M, Gunter M, Trethewey R, Kamlage B, Poorter H, Stitt M: Multilevel genomic analysis of the response of transcripts, enzyme activities and metabolites in Arabidopsis rosettes to a progressive decrease of temperature in the non-freezing range. Plant Cell Environ. 2008, 31 (4): 518-547. 10.1111/j.1365-3040.2007.01763.x.
Caldana C, Degenkolbe T, Cuadros-Inostroza A, Klie S, Sulpice R, Leisse A, Steinhauser D, Fernie AR, Willmitzer L, Hannah MA: High-density kinetic analysis of the metabolomic and transcriptomic response of Arabidopsis to eight environmental conditions. Plant J. 2011
Yun KY, Park MR, Mohanty B, Herath V, Xu F, Mauleon R, Wijaya E, Bajic VB, Bruskiewich R, de LRBG: Transcriptional regulatory network triggered by oxidative signals configures the early response mechanisms of japonica rice to chilling stress. BMC Plant Biol. 2010, 10: 16-10.1186/1471-2229-10-16.
Urano K, Maruyama K, Ogata Y, Morishita Y, Takeda M, Sakurai N, Suzuki H, Saito K, Shibata D, Kobayashi M, Yamaguchi-Shinozaki K, Shinozaki K: Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics. Plant J. 2009, 7: 1065-1078.
Cook D, Fowler S, Fiehn O, Thomashow MF: A prominent role for the CBF cold response pathway in configuring the low-temperature metabolome of Arabidopsis. Proc Natl Acad Sci USA. 2004, 101 (42): 15243-15248. 10.1073/pnas.0406069101.
Maruyama K, Takeda M, Kidokoro S, Yamada K, Sakuma Y, Urano K, Fujita M, Yoshiwara K, Matsukura S, Morishita Y, Sasaki R, Suzuki H, Saito K, Shibata D, Shinozaki K, Yamaguchi-Shinozaki K: Metabolic pathways involved in cold acclimation identified by integrated analysis of metabolites and transcripts regulated by DREB1A and DREB2A. Plant Physiol. 2009, 150 (4): 1972-1980. 10.1104/pp.109.135327.
Nakamichi N, Kusano M, Fukushima A, Kita M, Ito S, Yamashino T, Saito K, Sakakibara H, Mizuno T: Transcript profiling of an Arabidopsis PSEUDO RESPONSE REGULATOR arrhythmic triple mutant reveals a role for the circadian clock in cold stress response. Plant Cell Physiol. 2009, 50 (3): 447-462. 10.1093/pcp/pcp004.
Kaplan F, Kopka J, Haskell DW, Zhao W, Schiller KC, Gatzke N, Sung DY, Guy CL: Exploring the temperature-stress metabolome of Arabidopsis. Plant Physiol. 2004, 136 (4): 4159-4168. 10.1104/pp.104.052142.
Wienkoop S, Morgenthal K, Wolschin F, Scholz M, Selbig J, Weckwerth W: Integration of metabolomic and proteomic phenotypes: analysis of data covariance dissects starch and RFO metabolism from low and high temperature compensation response in Arabidopsis thaliana. Mol Cell Proteomics. 2008, 7 (9): 1725-1736. 10.1074/mcp.M700273-MCP200.
Salekdeh GH, Reynolds M, Bennett J, Boyer J: Conceptual framework for drought phenotyping during molecular breeding. Trends Plant Sci. 2009, 14 (9): 488-496. 10.1016/j.tplants.2009.07.007.
Armengaud P, Sulpice R, Miller AJ, Stitt M, Amtmann A, Gibon Y: Multilevel analysis of primary metabolism provides new insights into the role of potassium nutrition for glycolysis and nitrogen assimilation in Arabidopsis roots. Plant Physiol. 2009, 150 (2): 772-785. 10.1104/pp.108.133629.
Hu H, Dai M, Yao J, Xiao B, Li X, Zhang Q, Xiong L: Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proc Natl Acad Sci USA. 2006, 103 (35): 12987-12992. 10.1073/pnas.0604882103.
Xiao B, Huang Y, Tang N, Xiong L: Over-expression of a LEA gene in rice improves drought resistance under the field conditions. TAG Theoretical and applied genetics Theoretische und angewandte Genetik. 2007, 115 (1): 35-46.
Nelson DE, Repetti PP, Adams TR, Creelman RA, Wu J, Warner DC, Anstrom DC, Bensen RJ, Castiglioni PP, Donnarummo MG, Hinchey BS, Kumimoto RW, Maszle DR, Canales RD, Krolikowski KA, Dotson SB, Gutterson N, Ratcliffe OJ, Heard JE: Plant nuclear factor Y (NF-Y) B subunits confer drought tolerance and lead to improved corn yields on water-limited acres. Proc Natl Acad Sci USA. 2007, 104 (42): 16450-16455. 10.1073/pnas.0707193104.
Oh SJ, Kim YS, Kwon CW, Park HK, Jeong JS, Kim JK: Overexpression of the transcription factor AP37 in rice improves grain yield under drought conditions. Plant Physiol. 2009, 150 (3): 1368-1379. 10.1104/pp.109.137554.
Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, Amin N, Schwikowski B, Ideker T: Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13 (11): 2498-2504. 10.1101/gr.1239303.
Audenaert P, Van PT, Brondel F, Pickavet M, Demeester P, Van dPY, Michoel T: CyClus3D: a Cytoscape plugin for clustering network motifs in integrated networks. Bioinformatics. 2011, 27 (11): 1587-1588. 10.1093/bioinformatics/btr182.
Xia T, Hemert JV, Dickerson JA: OmicsAnalyzer: a Cytoscape plug-in suite for modeling omics data. Bioinformatics. 2010, 26 (23): 2995-2996. 10.1093/bioinformatics/btq583.
Schulze WX: Proteomics approaches to understand protein phosphorylation in pathway modulation. Curr Opin Plant Biol. 2010, 13 (3): 280-287.
Sugiyama N, Nakagami H, Mochida K, Daudi A, Tomita M, Shirasu K, Ishihama Y: Large-scale phosphorylation mapping reveals the extent of tyrosine phosphorylation in Arabidopsis. Mol Syst Biol. 2008, 4: 193-
Nakagami H, Sugiyama N, Mochida K, Daudi A, Yoshida Y, Toyoda T, Tomita M, Ishihama Y, Shirasu K: Large-scale comparative phosphoproteomics identifies conserved phosphorylation sites in plants. Plant Physiol. 2010, 153 (3): 1161-1174. 10.1104/pp.110.157347.
de la Fuente van Bentem S, Anrather D, Roitinger E, Djamei A, Hufnagl T, Barta A, Csaszar E, Dohnal I, Lecourieux D, Hirt H: Phosphoproteomics reveals extensive in vivo phosphorylation of Arabidopsis proteins involved in RNA metabolism. Nucleic Acids Res. 2006, 34 (11): 3267-3278. 10.1093/nar/gkl429.
Chen Y, Hoehenwarter W, Weckwerth W: Comparative analysis of phytohormone-responsive phosphoproteins in Arabidopsis thaliana using TiO2-phosphopeptide enrichment and mass accuracy precursor alignment. Plant J. 2010, 63 (1): 1-17.
Kline KG, Barrett-Wilt GA, Sussman MR: In planta changes in protein phosphorylation induced by the plant hormone abscisic acid. Proc Natl Acad Sci USA. 2010, 107 (36): 15986-15991. 10.1073/pnas.1007879107.
Zhang X, Yazaki J, Sundaresan A, Cokus S, Chan SW, Chen H, Henderson IR, Shinn P, Pellegrini M, Jacobsen SE, Ecker JR: Genome-wide high-resolution mapping and functional analysis of DNA methylation in arabidopsis. Cell. 2006, 126 (6): 1189-1201. 10.1016/j.cell.2006.08.003.
Gregory BD, Yazaki J, Ecker JR: Utilizing tiling microarrays for whole-genome analysis in plants. Plant J. 2008, 53 (4): 636-644.
Matsui A, Ishida J, Morosawa T, Mochizuki Y, Kaminuma E, Endo TA, Okamoto M, Nambara E, Nakajima M, Kawashima M, Satou M, Kim JM, Kobayashi N, Toyoda T, Shinozaki K, Seki M: Arabidopsis transcriptome analysis under drought, cold, high-salinity and ABA treatment conditions using a tiling array. Plant Cell Physiol. 2008, 49 (8): 1135-1149. 10.1093/pcp/pcn101.
Zeller G, Henz SR, Widmer CK, Sachsenberg T, Ratsch G, Weigel D, Laubinger S: Stress-induced changes in the Arabidopsis thaliana transcriptome analyzed using whole-genome tiling arrays. Plant J. 2009, 58 (6): 1068-1082. 10.1111/j.1365-313X.2009.03835.x.
Nie L, Wu G, Culley DE, Scholten JC, Zhang W: Integrative analysis of transcriptomic and proteomic data: challenges, solutions and applications. Crit Rev Biotechnol. 2007, 27 (2): 63-75. 10.1080/07388550701334212.
Bohmer M, Schroeder JI: Quantitative transcriptomic analysis of abscisic acid-induced and reactive oxygen species-dependent expression changes and proteomic profiling in Arabidopsis suspension cells. Plant J. 2011, 67 (1): 105-118. 10.1111/j.1365-313X.2011.04579.x.
Yang S, Vanderbeld B, Wan J, Huang Y: Narrowing down the targets: towards successful genetic engineering of drought-tolerant crops. Mol Plant. 2010, 3 (3): 469-490. 10.1093/mp/ssq016.
Reiland S, Finazzi G, Endler A, Willig A, Baerenfaller K, Grossmann J, Gerrits B, Rutishauser D, Gruissem W, Rochaix JD, Baginsky S: Comparative phosphoproteome profiling reveals a function of the STN8 kinase in fine-tuning of cyclic electron flow (CEF). Proc Natl Acad Sci USA. 2011.
Download references
Author information
Authors and affiliations.
Department of Biochemistry and Molecular Biology, Mail Stop 330, University of Nevada, Reno, Nevada, 89557, USA
Grant R Cramer
Gene Discovery Research Group, RIKEN Plant Science Center, 3-1-1 Koyadai, Tsukuba, 305-0074, Japan
Kaoru Urano & Kazuo Shinozaki
Univ. Bordeaux, ISVV, Ecophysiologie et Génomique Fonctionnelle de la Vigne, UMR 1287, Villenave d'Ornon, F-33882, France
Serge Delrot
Dipartimento di Biotecnologie, Università di Verona, Strada le Grazie 15, Verona, 37134, Italy
Mario Pezzotti
You can also search for this author in PubMed Google Scholar
Corresponding author
Correspondence to Grant R Cramer .
Additional information
Authors' contributions.
GRC contributed to all aspects of this manuscript. KU and KS contributed to the metabolomics and ABA signaling sections and Figure 2 . SD contributed to the introduction, ABA and sugar signaling sections. MP contributed to the time-series analyses section. All authors read and approved the final version of this manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Authors’ original file for figure 1
Authors’ original file for figure 2, rights and permissions.
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License ( http://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Reprints and permissions
About this article
Cite this article.
Cramer, G.R., Urano, K., Delrot, S. et al. Effects of abiotic stress on plants: a systems biology perspective. BMC Plant Biol 11 , 163 (2011). https://doi.org/10.1186/1471-2229-11-163
Download citation
Received : 05 September 2011
Accepted : 17 November 2011
Published : 17 November 2011
DOI : https://doi.org/10.1186/1471-2229-11-163
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
- Abiotic Stress
- Reactive Nitrogen Species
- Antisense Transcript
- Abiotic Stress Response
BMC Plant Biology
ISSN: 1471-2229
- General enquiries: [email protected]
An official website of the United States government
Official websites use .gov A .gov website belongs to an official government organization in the United States.
Secure .gov websites use HTTPS A lock ( Lock Locked padlock icon ) or https:// means you've safely connected to the .gov website. Share sensitive information only on official, secure websites.
- Publications
- Account settings
- Advanced Search
- Journal List
Abiotic Stress in Crop Species: Improving Tolerance by Applying Plant Metabolites
Francisca godoy, karina olivos-hernández, claudia stange, michael handford.
- Author information
- Article notes
- Copyright and License information
Correspondence: [email protected] ; Tel.: +56-2-2978-7263
These authors contributed equally to this work.
Received 2020 Oct 30; Accepted 2020 Dec 4; Collection date 2021 Feb.
Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/ ).
Reductions in crop yields brought about by abiotic stress are expected to increase as climate change, and other factors, generate harsher environmental conditions in regions traditionally used for cultivation. Although breeding and genetically modified and edited organisms have generated many varieties with greater abiotic stress tolerance, their practical use depends on lengthy processes, such as biological cycles and legal aspects. On the other hand, a non-genetic approach to improve crop yield in stress conditions involves the exogenous application of natural compounds, including plant metabolites. In this review, we examine the recent literature related to the application of different natural primary (proline, l -tryptophan, glutathione, and citric acid) and secondary (polyols, ascorbic acid, lipoic acid, glycine betaine, α-tocopherol, and melatonin) plant metabolites in improving tolerance to abiotic stress. We focus on drought, saline, heavy metal, and temperature as environmental parameters that are forecast to become more extreme or frequent as the climate continues to alter. The benefits of such applications are often evaluated by measuring their effects on metabolic, biochemical, and morphological parameters in a variety of crop plants, which usually result in improved yields when applied in greenhouse conditions or in the field. As this strategy has proven to be an effective way to raise plant tolerance to abiotic stress, we also discuss the prospect of its widespread implementation in the short term.
Keywords: drought stress, heavy metal stress, primary metabolite, salt stress, secondary metabolites
1. Introduction
Abiotic stress is one of the most important problems currently faced by agriculture. It causes serious losses in crop production worldwide and reduces planted acreage. Amidst a growing population and climate change, this scenario becomes increasingly complex. Because the world population is forecast to increase from 7 to 9–10 billion people by 2050, an increase of between 60 and 110% in global food production will be required ([ 1 ] and references therein). Arable lands are also affected by migration to cities; as urban areas expand, they encroach more into surrounding, often fertile land, which is another factor that pushes agriculture into areas that are less-suited to crop cultivation [ 2 ]. Due to deforestation and the excessive use of fossil fuels, atmospheric CO 2 levels have increased from 280 to 400 ppm and are expected to rise to 800 ppm by 2100 [ 3 , 4 ]. Furthermore, climate change causes extreme weather, such as brusque temperature fluctuations, extreme precipitation, and drought, among other effects. This is subjecting crop species to increased abiotic stress and impacts production. For example, 45% of arable lands are subjected to drought [ 5 ], and the world area under aridity stress increased from 17 to 27% between 1950 and 2000 [ 6 ]. Furthermore, salinity accounts for considerable decreases in crop productivity [ 7 ], because most crop species are sensitive to salt stress (1.0–1.8 dS m −1 ), and it negatively impacts yield by 10 to 50%, depending on the salt concentration present [ 8 ]. Anthropogenic activities, such as mineral extraction and the over-application of fertilisers and pesticides, are also related to increases in heavy metal pollution, including excess hexachromium (Cr), cadmium (Cd), arsenic (As), lead (Pb), copper (Cu), and mercury (Hg). These pollutants affect plant developmental processes, such as seed germination, and the rates of photosynthesis, respiration, and transpiration, thus reducing growth, yield and quality [ 9 , 10 ] ( Figure 1 ). These situations thus demand changes in agricultural practices to respond to the negative impacts of climate change and anthropogenic activities.

Abiotic stress reduces crop yield. Environmental stress factors, such as heat, cold, drought, salinity, and the presence of heavy metals such cadmium, copper, and chromium, elicit stress responses in plants, including an accumulation of reactive oxygen species (ROS) and reduced photosynthetic activity, which ultimately lower plant growth and thus crop yields.
Plants are commonly subjected to different kinds of abiotic stress, such as UV, high temperatures, drought, floods, and heavy metals. As sessile organisms, they have evolved structural and metabolic adaptations to survive these conditions, such as increased root area and leaf curling when exposed to drought; greater production of antioxidant compounds such as carotenoids, proline, and ascorbic acid; and higher activity of enzymes related to the scavenging of reactive oxygen species (ROS). Abiotic stress causes an imbalance of pro-oxidant and antioxidant compounds, also known as oxidative stress, via processes that have been covered by excellent reviews [ 11 ], including in crop species [ 12 ]. Briefly, abiotic stress factors frequently favours stomatal closure, increasing the activity of the photorespiratory pathway, and triggering the production of hydrogen peroxide (H 2 O 2 ). Additionally, the diminished uptake and assimilation of CO 2 results in an imbalance of electron flow through the photosynthetic electron transport chain, again sparking the production of superoxide radicals and singlet oxygen ( 1 O 2 ). Together with hydroxyl radicals (OH • ), the excess H 2 O 2 , superoxide radicals and 1 O 2 are highly reactive to proteins, lipids, and nucleic acids, damaging them and causing cell death. A common marker for lipid peroxidation, considered one of the most damaging processes, is malondialdehyde (MDA), which is an oxidised product of membrane lipids. To overcome oxidative stress, plants possess enzymatic and non-enzymatic detoxification systems. In the enzymatic system, the enzymes superoxide dismutase (SOD), catalase (CAT), monodehydroascorbate reductase (MDHAR), dehydroascorbate reductase (DHAR), glutathione reductase (GR), glutathione transferase (GST), ascorbate peroxidase (APX), glutathione peroxidase (GPX), and guaiacol peroxidases (POXs) among others, are involved in ROS scavenging. These enzymes present higher activity when plants are subjected to stress. The non-enzymatic mechanism involves the production of compounds such as proline, glutathione (GSH), ascorbic acid, carotenoids, flavonoids, and tocopherols, that alleviate oxidative damage by neutralising ROS [ 13 , 14 ]. In the case of GSH, not only its concentration but the ratio with its oxidised form, glutathione disulphide (GSSG), is important. Crop varieties presenting higher endogenous amounts of some of these compounds display improved tolerance. However, under high intensity or continued stress, these strategies are not enough to alleviate the damage, and plant growth and yield are severely affected.
Currently there are several strategies to increase abiotic stress tolerance in crop species: (1) Genetically engineered organisms (GMOs), including transgenic, cisgenic, and intragenic crops; (2) traditional breeding programs; (3) new breeding technologies (NBTs), including the use of the Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR/Cas) edition strategy; and (4) the application of plant biostimulants. Biostimulants are defined as plant fertilisers that improve nutrient use efficiency, tolerance to abiotic stress, quality traits or availability of confined nutrients in the soil or rhizosphere [ 15 ].
Although many GMOs have been bioengineered to tolerate abiotic stress, their practical use depends on the politics and legal aspects of each country, and it is a lengthy process before they are accepted for cultivation and commercialisation. Although more than 500 transgenic crops have been approved for cultivation, only 12 are related to tolerance to abiotic stress due to the high complexity of this trait [ 16 ]. Furthermore, there is a generally negative public perception regarding transgenic crops and a lengthy regulatory process to obtain permission for cultivation. It has been estimated that, on average, it costs USD 35 million to process the regulatory safety assessments and authorisations, and that it takes approximately 13 years from the development of a transgenic crop to its commercial launch [ 16 , 17 ]. Regarding genetically edited crops, the CRISPR mushroom was the first edited commercialised food [ 18 ], but their governance is still being developed in many countries [ 19 ]. Crop varieties obtained by conventional breeding programs are not limited by environmental and biosafety regulations, but present the problem that it takes ten to twenty years to develop a new variety, depending on the species [ 20 ].
On the other hand, a non-genetic approach to improve crop yield in stress conditions involves the exogenous application of biostimulants, including naturally occurring plants and fungal and microbial metabolites. For example, the application of amino acids, sugars, secondary metabolites, or artificial compounds has proven to be an effective way to increase plant tolerance to stress, and is relatively easy to implement in the short term. It has been reported that the application of endogenous metabolites enhances the plant stress response, and these compounds have the advantage in that they participate in combating different kinds of stress. Phytohormones, as key regulators of plant development, have also been applied to crops in order to improve tolerance to different kinds of abiotic stress. Exogenous application of auxin, cytokinins, abscisic acid, salicylic acid, and gibberellin have all proven useful in alleviating the effects of abiotic stress, as have transgenic means to enhance endogenous hormone production ([ 21 , 22 , 23 , 24 , 25 ] and references therein). As the role of phytohormones in abiotic stress has been widely described by other colleagues in the aforementioned publications, this review will focus on primary and secondary metabolites.
It is known that in plants, fungi, and microorganisms, more than 200,000 substances are produced, and they can be grouped into primary and secondary metabolites [ 26 ]. Primary metabolites are synthesised by all organisms and are essential for central, vital processes such as growth and development, and include proteins, lipids, and carbohydrates. In plants, secondary metabolites are synthesised in different cellular compartments, and include phenolics, terpenoids, and alkaloids as main groups, whose many functions include protection against abiotic and biotic stress. Different metabolic routes are involved in the synthesis of these molecules, such as the shikimate pathway, the acetate–malonate pathway, and side reactions emanating from glycolysis and the tricarboxylic acid (TCA) cycle [ 26 , 27 , 28 ]. Compared with primary metabolites, secondary metabolites are produced in low levels (<1% dry weight) and are specific to certain organs or developmental stages.
In this review, we present the latest literature on how the exogenous application of key natural primary and secondary metabolites has been used to improve abiotic stress tolerance (salinity, drought, and heavy metal) in both greenhouse and field conditions, with a focus on their effects on growth, physiological parameters, and yield in crop species. Exogenous metabolite application is an easy-and-feasible-to-implement strategy for producers, although the time interval in which they exert their beneficial effects depends on dose, the time of application and crop species. Finally, we comment on the challenges to overcome so that growers and producers can profit from the knowledge generated in this fast-developing area.
2. Primary Metabolites
2.1. proline.
Proline is one of the most studied osmoprotectants in plants. In 1954, Kemble and McPherson described proline accumulation in wilting rye grass ( Lolium perenne ), and since then, it has been the focus of many studies. Proline can function as a molecular chaperone, protecting protein integrity. Moreover, it acts as a singlet oxygen quencher, contributing to the maintenance of redox balance [ 29 , 30 ]. However, the beneficial effects observed in abiotic stress tolerance depend on species, concentration, phenological time of application, and application system.
Extensive literature exists on the beneficial effects of proline when applied to crop species under abiotic stress. For example, Teh et al. (2016) [ 31 ] applied proline (5 and 10 mM) to 30-day-old rice ( Oryza sativa ) plants cultivated in vitro under saline stress; the higher concentration significantly increased plant height and shoot and root number when compared to control stressed plants. Additionally, in rice, Tabssum et al. (2019) [ 32 ] observed improved water relations in salt+proline-treated plants grown in greenhouse conditions, as well as an increase in SOD and CAT activities, indicating an enhanced effect of proline on ROS scavenging when compared to salt-stress conditions. However, 50 mM-proline treatments had adverse effects on bean ( Vicia faba ), where plant growth and photosynthetic pigment accumulation were similar to that of the sea water treatments [ 33 ], indicating toxic effects. Thus, dose optimisation is a crucial step towards its use for yield improvement in agriculture.
It has been reported that proline treatment in the context of abiotic stress induces changes at the structural and ultrastructural levels. An increase in root surface is one of the strategies adopted by plants to overcome nutrient or water shortage. Proline application during salt stress induced changes in root surface in rice [ 31 ], increasing the number of roots per plant. Regarding the aerial part, proline application in salt-stressed plants produced structural changes in stems and leaves. Rady et al. reported an increase in cortical layers, xylem vessel diameter, and cortex thickness in two varieties of lupine ( Lupinus termis ) grown in salinity conditions, when 3, 6 or 9 mM proline was foliar-applied at 20, 35, and 50 days after sowing, and the best results were obtained with a proline concentration of 6 mM [ 34 ]. At the ultrastructural level, treatments with proline in rice leaves under control conditions produced swelling of thylakoids and an increase in total starch area. Salt stress also had this effect, especially 100 mM NaCl. However, when proline was applied in rice plants under saline stress, chloroplasts maintained their structural integrity, and no thylakoid swelling was observed. This is consistent with reports describing a protecting role of proline on thylakoid membranes by scavenging ROS [ 35 ].
The effectiveness of proline in the alleviation of metal toxicity has also been studied. Treatment by excess caused a decrease in growth parameters, an increase in MDA and H 2 O 2 levels, and SOD and POX activities in eggplant ( Solanum melongena ). However, As+proline treatment caused an increment in dry weight (DW) and root length, reduced As content in eggplant seedlings, decreased H 2 O 2 and MDA levels, and increased activity of ROS scavenging-related enzymes. It also generated higher activity levels of the two proline biosynthesis enzymes, Δ1-pyrroline-5-carboxylate synthetase (P5CS) and proline dehydrogenase [ 36 ]. Likewise, in rice treated with Cr, proline application significantly decreased MDA levels [ 37 ]. In a pot experiment where wheat ( Triticum aestivum ) was subjected to Cu stress, proline application increased yield (100-grain weight), although it depended on the cultivar used [ 38 ].
Further studies evaluating crop yield improvement under field conditions and long-term stress are needed in order to validate the use of proline by producers.
2.2. L-Tryptophan (TRP)
Auxin is the most studied hormone in plants. Because abiotic stress impairs plant development and growth, one of the approaches to overcome this has been the application of L-tryptophan (β-3-indolylalanine), an essential amino acid that serves as precursor for auxin biosynthesis [ 39 ].
The effect of TRP in ameliorating salt-induced damage in crops has also been studied. Red peppers ( Capsicum annuum ) grown in pots in a wire house were subjected to different salinity levels and TRP treatments. Plant growth parameters, such as plant height, root length, shoot, and root DW, increased in all conditions when TRP was applied, as did chlorophyll and carotenoid levels. Moreover, fruit weight was higher when compared to untreated controls. At high salinity, control plants did not produce fruit; however, under TRP treatment, plants were able to develop fruits, albeit small ones [ 40 ]. Other studies carried out in sugar beet ( Beta vulgaris ) showed that several root parameters at harvest were improved in plants subjected to different salinity levels whose seeds had been pre-soaked with TRP when compared to salt-stressed plants which had not been treated with this amino acid [ 41 ]. Furthermore, salinity decreased root quality by increasing K + , Na + , and α-amino-N, which are considered impurity parameters. TRP seed treatment improved said parameters, thus enhancing root quality [ 41 ].
Regarding drought stress, TRP has proven to improve yield. Jamil, 2018 [ 42 ], performed a pot experiment, where wheat ( Triticum aestivum ) was subjected to different irrigation treatments. TRP was foliar sprayed every week, starting 15 days after germination and ending two weeks after the filling stage. TRP treatment increased growth parameters, such as plant height, root length, and root DW, in all irrigation conditions when compared to control plants, as well as the amount of photosynthetic pigments. TRP also increased yield, either under control or drought conditions [ 42 ].
Moreover, TRP improves yield in crops subjected to heavy metal stress. In rice subjected to Cd stress, a decrease in plant growth (plant height and number of tillers) and yield parameters (number of panicles, 1000 grain weight, and paddy yield) was observed when compared to control plants. However, Cd+TRP treatment increased those parameters when compared to plants subjected to Cd stress alone [ 43 ], indicating that TRP is effective at alleviating Cd-induced damage.
Hanci et al. evaluated the effect of TRP on seed germination under temperature stress [ 44 ]. They treated onion ( Allium cepa ) and leek ( Allium porrum ) seeds with different concentrations of TRP, and subjected them to 7, 21, and 35 °C in a germination chamber for 21 days. In onion, germination percentage at the lowest temperature was slightly increased by TRP treatment (125 ppm), although higher doses of this metabolite decreased the germination rate. In leek, beneficial effects of TRP were observed only in cold stress conditions (7 °C). The authors conclude that although TRP application may improve germination, further studies are needed to identify the appropriate dose for each species [ 44 ].
In summary, TRP has been seen to induce plant growth and yield parameters in crops subjected to different kinds of stress, such as salt, drought, and excess Cd, although the cost of applying in the field must be considered and evaluated.
2.3. Glutathione (GSH)
GSH (γ-l-glutamyl-l-cysteinylglycine) is a ubiquitous low molecular weight tripeptide, composed of the essential amino acids glutamine, cysteine, and glycine [ 45 ], although some plants exhibit a variation called homoglutathione, with the same biological properties. It is present in most plant tissues, and among its major functions are ROS detoxification, formation of phytochelatins that bind heavy metals, detoxification of methylglyoxal, and its role as a cysteine reservoir [ 46 , 47 ].
GSH application in mungbean ( Vigna radiata ) seedlings grown under salt stress in growth chambers improved the relative water content (RWC) and chlorophyll ( a and b ) levels at 24 and 48 h after GSH treatment. Classic stress marker levels such as H 2 O 2 , MDA, and superoxide generation rate were also significantly decreased in the salt+GSH treatment when compared to the salt stress treatment alone. The GSH/GSSG ratio was also significantly increased when comparing GSH treatment to salt stress alone, indicating a better redox state. Enzyme activities from the ROS scavenging system (SOD, APX, GST, MDHAR, DHAR, GR, and GPX) were also increased in plants subjected to salt+GSH when compared to control plants under salt stress alone [ 48 ]. Zhou et al. also evaluated the enzyme activity and transcript abundance of ROS scavengers under GSH and salt treatments in tomatoes ( Solanum lycopersicum ) grown under greenhouse conditions [ 49 ]. They found that salt stress decreased the activity levels of SOD, POD, CAT, MDHAR, DHAR, and APX, as well as GSH, when compared to control conditions, but GSH treatment improved enzyme activity when compared to salt stress. The foliar application of GSH to tomato seedlings grown under salt stress reduced H 2 O 2 , MDA, and superoxide generation rate levels, as well as a decrease in Na + and Cl - uptake and accumulation when compared to salt-stressed plants [ 49 ]. Even though the study evaluated the effect of GSH at 5, 10, and 15 days after treatments, the results were consistent with those reported for other species [ 48 , 49 ].
Moreover, eleven soybean ( Glycine max ) varieties grown under nets showed significant increments in plant growth and production parameters when GSH was applied to leaves of salt-stressed plants in comparison to stress-only conditions [ 50 ]. For example, the number of seeds per plant increased by 22–62% (depending on the genotype examined) when compared to stressed treatment. Furthermore, other parameters interesting for their impact on crop yield were significantly improved, such as pods per plant (by 12–60%) and yield per plant (by 16–67%), when compared to plants subjected to salt stress alone [ 50 ]. Finally, genotypes categorised as susceptible to salt stress presented better responses when also treated with GSH [ 50 ], emphasising the importance of conducting studies of the effects of exogenous metabolite application for each genotype. Such studies will be very important for encouraging the cultivation of salt-susceptible varieties with desirable or unique commercial traits, even in saline soil conditions.
Given the importance of GSH as a precursor for phytochelatin synthesis, its effect on alleviating heavy metal toxicity in crops has been evaluated. Khan et al. reported that Pb toxicity decreased photosynthetic attributes such as photosynthetic rate, stomatal conductance, intercellular CO 2 concentration, transpiration rate, and photosynthetic pigments, and also increased MDA, H 2 O 2 , and Pb levels in upland cotton ( Gossypium hirsutum ) grown in hydroponic conditions [ 51 ]. GSH applied to the nutrient medium improved the aforementioned parameters when compared to Pb-treated plants, reaching in many cases the levels present in control conditions. It also amended Pb-induced damage observed at the ultrastructural level, maintaining the integrity of chloroplasts and other organelles [ 51 ]. Additionally, in a study conducted by Kim et al. [ 52 ], GSH improved seed germination and seedling growth in tobacco ( Nicotiana tabacum ), red pepper and even a model Brassicaceae species ( Arabidopsis thaliana ) under Hg stress. They also demonstrated that GSH presented stronger binding affinity to Hg when compared to other heavy metals such as Cd, Cu, or zinc [ 52 ], indicating that GSH may be better suited to alleviate Hg-induced damage.
GSH application also had positive effects on plants subjected to high temperatures. Mungbean seedlings grown in growth chambers were exposed to 42 °C or normal conditions (25 °C) and pretreated with GSH 24 h before the high temperature treatment. Leaf RWC, chlorophyll, and proline content increased in plants treated with GSH when compared to the plants subjected to high temperature. The stress markers MDA and H 2 O 2 content, as well as superoxide generation rate, were diminished in GSH-treated plants when compared to the stress treatment. Additionally, the ascorbic acid content and GSH/GSSG ratio were increased in GSH-treated plants in comparison to the levels found in heat-stressed plants [ 53 ].
2.4. Citric Acid (CA)
Citric acid (CA), the initial intermediate of the TCA cycle, is synthesised by citrate synthase from the condensation of oxaloacetate and acetyl CoA [ 54 ]. This molecule is a mild antioxidant that can also quelate metals such as Cu, Pb, and aluminium [ 55 ]. Problems induced by salinity can be alleviated by CA application. For instance, the addition of CA to the irrigation solution of tomatoes in calcareous soils (high concentration of CaCO 3 ) improved the uptake and assimilation of Zn, Na, Ca, and N in leaves, and Mn, Na, Mg, and P in fruits [ 56 ]. Likewise, maize ( Zea mays ) plants grown in soil with high salinity and 100–200 ppm CA showed restored height, chlorophyll a and b contents, enhanced growth, and an improvement in yield when compared to salt-treated controls [ 57 ].
Regarding heavy metal stress, exogenous applications of CA to mustard ( Brassica juncea ) leaves reduced the harmful effects of Cd by improving the enzymatic antioxidant response, via increased CAT, POX, and SOD activities. Additionally, the chlorophyll content and stomatal conductance and aperture all rose, leading to a 14% increase in CO 2 availability, improved photosynthetic rate (by 42%) and growth (by 12%) of the Cd+CA plants compared to mustard stressed with Cd [ 58 ].
On other hand, the application of CA can revert the reduction in germination, mineral absorption, hormone homeostasis, and lower growth and yield produced by Pb stress. In the case of castor bean ( Ricinus communis ) grown in soil with 600 mg kg −1 Pb and with applications of 5 mM CA, an improvement was observed in the length of the roots and shoots and the number and area of leaves per plant compared to the plants grown in soil with Pb alone, resulting in phenotypes similar to those of control plants (without application of Pb or CA). The photosynthetic parameters, transpiration rate, water use efficiency, and stomatal conductance were improved, as were chlorophyll a and b and carotenoid levels. The activities of antioxidant enzymes, such as POX, CAT, APX, and SOD, were also enhanced in both roots and leaves of Pb+CA plants [ 59 ].
Another case is the stress inflicted by Cu, a metal cofactor for many enzymes involved in electron transfer and an essential micronutrient. Nevertheless, at higher concentrations, it has a toxic effect on seed germination, photosynthetic activity, and cell division; induces changes in the ionic form of nutrients; and generates oxidative stress. It has been shown that canola ( Brassica napus ) can tolerate soils contaminated with an excess of Cu, but plant growth, shoot and root length, and photosynthetic pigments all suffer decreases, whilst the activity of antioxidant enzymes increases, affecting yield. However, the application of CA can counteract this effect [ 60 ]. Specifically, the application of 2.5 mM CA in soil together with 100 µM Cu recovered growth and increases of 20–31% were observed in shoot and root length and the number and area of leaves when compared with Cu (100 µM) alone. Similarly, the photosynthetic pigment content and the activities of antioxidant enzymes (SOD, CAT, POX) were raised in Cu+CA-treated canola compared to the Cu-treated controls. A similar finding was discovered in pea ( Pisum sativum ), in that the application of 100 µM CA in the Cu-containing germination medium restored biomass production in roots and shoots, reduced cell death, and corrected cellular redox state versus seeds treated with Cu alone [ 61 ]. In white jute ( Corchorus capsularis ), a plant that is used in phytoremediation due to its tolerance to heavy metals, the application of 2 mM CA in the solution media improved parameters like plant growth, biomass, chlorophyll, and carotenoid contents, whilst lowering the activities of SOD and POX [ 62 ].
Cr, another heavy metal, can become toxic to plants in high concentrations. In plants, the stress induced by Cr diminishes seed germination, growth, uptake of mineral nutrients available in the soil, and photosynthesis. Such effects can be reverted by the exogenous application of CA. In rice plants, supplementing the irrigation solution with 100 µM CA restored parameters like root and shoot length and DW. Rice plants also had higher internal Cr concentrations (by 50%), demonstrating the capacity of CA to chelate metals, mirrored by enhanced activities of the detoxification enzymes (CAT, POD, SOD, and GR) and metabolites (GSH and proline), enabling the Cr+CA plants to cope better with scavenging ROS [ 63 ]. Similar findings have also been discovered in Cr+CA-treated spinach ( Spinacia oleracea [ 64 ]).
The multiple effects of adding the aforementioned primary metabolites are summarised in Figure 2 , and the principles of their modes of action also hold true for studies that have evaluated the consequences of applying secondary metabolites to different crop plants in greenhouse and field conditions.

Application of primary and secondary plant metabolites ameliorates the negative effects of abiotic stress. By applying primary metabolites such as proline, tryptophan (TRP), glutathione (GSH), and citric acid (CA), or secondary metabolites like polyols, lipoic acid (LA), ascorbic acid (AA), glycine betaine (GB), α-tocopherol (α-Toc), and melatonin as foliar sprays or in irrigation water, the tolerance of crops is enhanced when faced with environmental challenges.
3. Secondary Metabolites
3.1. polyols.
Polyols or sugar alcohols are organic compounds derived from sugars, in which the carbonyl group (CO) has been reduced to an alcohol (OH). They include linear and cyclic forms, termed alditols and inositols, respectively. Examples of polyols are mannitol (reduced form of mannose), sorbitol (from glucose), and inositol, synthesised from glucose 6-phosphate followed by dephosphorylation [ 65 , 66 , 67 ].
3.1.1. Mannitol
Protective effects on maize with foliar applications of mannitol (15 and 30 mM) have been observed under saline stress (100 mM NaCl). The combined treatment increased total plant DW, RWC, and chlorophyll content ( a and b ), demonstrating that mannitol was mitigating, directly and/or indirectly, the harmful effects of saline stress and enhanced biomass production due to its role in cellular osmotic adjustment [ 68 ]. Such beneficial properties have also been discerned in the field: the reduced productivity seen in saline-stressed cabbage ( Brassica oleracea ) was alleviated by the foliar application of 2–4 g L −1 mannitol [ 69 ].
As mentioned above, Cr stress affects several important parameters of crops. The application of mannitol to pot-grown maize can alleviate these effects. The height, leaf area, root length, number of leaves per plant, and DW of all plant parts were improved by the foliar application of 50 and 100 mg L −1 mannitol to cultivars under Cr stress (5 and 10 mg kg −1 soil) when compared to plants without mannitol, and phenotypes were similar to control plants (without Cr or mannitol). Oxidative stress levels in roots and leaves were also lowered, especially in a more-tolerant cultivar, as well as the activity of antioxidant enzymes (SOD, POD, CAT, and APX) and the content of chlorophylls and carotenoids, suggesting a protective role of mannitol in maize under heavy metal stress [ 70 ]. Even though the application of 100 mM mannitol alone might have negative effects on wheat growth under normal conditions, because of induction of stomatal closure, when this polyol was combined with Cr (0.25 and 0.5 mM), pot-grown plants had enhanced shoot and root DWs, as well as husk and kernel weights, facilitated by increases in antioxidant systems in both leaves and roots [ 71 ].
Boron (B) toxicity is another kind of stress that has been observed mainly in arid and semi-arid environments, such as those found in South Australia, Turkey, California, and Chile. At high concentrations, B causes necrotic lesions in leaf tips and patches on leaves, reducing photosynthesis, growth and yield [ 72 ]. Two wheat cultivars, with different sensitivities to B, were analysed under increasing B concentrations (0, 30, 45, and 60 mg kg −1 ) and mannitol treatments (0, 1, 5, and 10 g kg −1 ) in pots. It was observed that the content of B in leaves was higher in the sensitive cultivar, and that application of mannitol reduced considerably the B content in both cultivars. In this crop, it was determined that 1 g kg −1 mannitol was more effective than the other concentrations in reducing the symptoms of B toxicity [ 73 ].
3.1.2. Sorbitol
The eggplant is a crop classified as moderately sensitive to salinity. To mitigate the damage caused by salt, the effects of foliar applications of sorbitol (5 and 10 mg L −1 ) to plants in greenhouse conditions subjected to different NaCl treatments (1.5, 3, and 6 dS m −1 ) was studied. It was observed that 5 mg L −1 sorbitol was the most efficient, increasing parameters like plant height, the number, weight, and diameter of fruits, as well as chlorophyll a and b and carotenoid levels in all salt treatments, compared to the controls (without sorbitol) [ 74 ]. Studies in rice [ 75 ] and wheat [ 76 ] have uncovered similar effects, although finding the correct concentration is key for success. For example, in rice, 10 mM sorbitol lowered H 2 O 2 and MDA levels in 170 mM NaCl-treated plants, increasing growth and DW [ 75 ], whereas in wheat, either 50 or 100 mM sorbitol had particularly beneficial effects on root and shoot lengths and DW, proline content, and photosynthetic pigment levels [ 76 ].
3.1.3. Inositol
The application of this cyclic polyol has been used to improve abiotic stress tolerance in a few cases. For instance, pre-treating crab apples ( Malus hupehensis ) with 50 µM myo-inositol in the hydroponic media reduced salt stress (200 mM) symptoms, as determined by improvements in plant height and DW, as well as root length, volume and surface area, compared to plants subjected to salt stress, whilst lowering relative electron leakage, H 2 O 2 production, and Na+ accumulation [ 77 ].
In the case of drought stress, exogenous myo-inositol can reduce its harmful effects by protecting the water status of leaves. It was observed that foliar application of 5, 15, and 25 µM myo-inositol to pepper plants under drought stress (7 days without watering) in pots raised the RWC and reduced H 2 O 2 , proline and MDA levels [ 78 ].
3.2. Lipoic Acid (LA)
Lipoic acid (LA, 6,8-thioctic acid or 1,2-dithiolane-3-pentanoic acid) is a powerful sulphur-containing antioxidant, whilst also acting as a cofactor in several multienzyme complexes involved in primary metabolism [ 79 ].
The antioxidant properties of LA have been used to benefit crop production. For example, in canola under salinity stress (<150 mM NaCl), the foliar application of 100 µM LA reduced lipid peroxidation levels, and raised cysteine content and POD and CAT activities, improving overall yield and providing tolerance to salt stress [ 80 ]. Additionally, in pot-grown wheat irrigated with different dilutions of seawater (<14.6 dS m −1 ), the foliar application of 0.1 mM LA buffered the deleterious effects by increasing levels of proline, the RWC, and activities of antioxidant enzymes (such as CAT, SOD, POX, and APX), which benefited agronomic parameters (leaf area, plant height, and grain yield) and improved tolerance to salt stress [ 81 ].
In the case of drought stress, exogenous application of LA also can be beneficial to plants, as shown in hydroponically propagated maize. Supplementation of the nutrient solution with PEG6000 simulates water deficit. However, the addition of 12 µM LA in the growth media can reduce lipid peroxidation and increase RWC, endogenous LA, chlorophyll a and b levels, and the expression of key genes ( rubisco large and small subunits ( LSU and SSU ), whilst also improving stomatal conductance and photosynthetic and transpiration rates compared to plants treated with PEG6000 alone [ 82 ].
Similar effects occur in Pb-treated (1.5 mM) wheat seeds, imbibed with 2 µM LA; these had longer roots and coleoptiles, even though both organs contained more Pb than seeds not treated with LA. The altered ratio of GSH/GSSG demonstrated that the LA-treated plants had a greater capability to withstand the potentially toxic effects of internal Pb [ 83 ].
These examples of the use of LA clearly highlight how the combination of species and abiotic stress influences the dose of the metabolite required to alleviate negative effects (100 µM canola/saline; 12 µM maize/drought; 2 µM wheat/Pb).
3.3. Ascorbic Acid (AA)
Ascorbic acid (AA), or vitamin C, is another secondary metabolite which can be synthesised de novo from several pathways, such as D-glucose, L-galactose, uronic acid, L-gulose, and myo-inositol pathways, or by recycling its oxidised form. It is a cofactor for many enzymes, and can neutralise ROS, repair oxidised organic molecules, and regulate physiological processes (such as cell division, growth, development, and stress tolerance) in plants [ 84 , 85 , 86 ].
The application of this metabolite can improve the tolerance of many species to a variety of abiotic stresses. In the cereal crop, quinoa ( Chenopodium quinoa ), different intensities of drought stress reduce quinoa shoot and root length, DW, levels of chlorophylls a and b , and total carotenoids, whilst raising H 2 O 2 and soluble sugar levels. However, when 150 mg L −1 AA was applied to leaves during the vegetative stage in greenhouse conditions, tolerance to drought stress was dramatically enhanced, as demonstrated by improvements in metabolic, biochemical, morphological and production parameters [ 87 ]. Similarly, for two pot-grown varieties of peach tree ( Prunus persica ), growth, nutrient uptake, yield, and fruit quality were all favoured by spraying leaves with 250 ppm AA subjected to water stress [ 88 ], as a consequence of higher net CO 2 assimilation rate, stomatal conductance, proline levels, and antioxidant enzyme activities. Such positive effects have also been seen in the field: the application of AA to wheat leaves (200 mg L −1 ) over two seasons in an experimental farm increased the relative water and chlorophyll content, with higher antioxidant enzyme activities (CAT and POX), resulting in improvements in the number of grains per spike, the number of spikes, and, consequently, yield [ 89 ].
In wheat under stress by Pb (2 mM), the application of 0.6 mM AA in the watering solution restored parameters such as root and shoot length; fresh weight (FW); the content of N, P, K, Ca, Mg, and cysteine; and total chlorophyll content, allowing the plant to develop with no serious consequences on yield, thus resembling treatment controls (without Pb or AA) and performing better than Pb-treatment alone. The antioxidant effect of AA also protected the chloroplasts from oxidative damage, as well as altering the concentration of plant hormones such as auxin, gibberellins, and abscisic acid [ 90 ].
Heat is another stress that affects the defence system of plants. For example, cotton is cultivated in various climates, but is sensitive to extreme temperatures, although even minor variations in the temperature can reduce the photosynthetic rate and generate ROS. However, the foliar spray application of 70 ppm AA to cotton cv. AA-802 can enhance the activity of SOD and CAT, raise chlorophyll content and antioxidants, and alleviate oxidative stress to such an extent that fibre lengths and, thus, the yields of the plants are affected to a much lesser extent or may even improve compared to the controls [ 91 ]. Such results are similar to those discovered in a previous study [ 92 ] in which the application of different concentrations of AA to cotton plants during the squaring and flowering stages under heat stress enhanced their RWC, total chlorophyll, the activity of antioxidant enzymes (POX, CAT, and SOD), and the yield compared to controls sprayed with water.
At the other extreme, low temperatures (chilling stress) can also affect crops, causing phenotypic changes such as chlorosis, wilting, metabolic changes, and the leakage of cellular solutes. An example of this can be seen in tomato plants stressed by chilling (4 °C), where leaf area, length, and width are reduced [ 93 ]. However, chilled seedlings incubated in AA solution (0.5 mM for 48 h) behaved similarly to the non-chilled controls, showing reduced oxidative damage and increased proline, chlorophyll, mineral nutrition, and expression of molecular chaperones (HSP70, HSP90, and HSP80) and CAT genes [ 93 ]. Likewise, AA application can also aid fruits, even after harvesting. In banana ( Musa spp), chilling affects many phenotypical characteristics important for consumers, such as the colour of fruit peel and pulp, that hinder the sale of the product. Yet a simple procedure of immersing the fruits in a solution of AA at different concentrations (the best was 9 mM) followed by cold storage (27 days at 6 °C) reduced the overall chilling injury index score, reflected in improvements in total chlorophyll, total phenolic and flavonoid content, and the activity of antioxidant enzymes (APX, POD, SOD, and CAT) [ 94 ]. AA applications can even improve extreme chilling (freezing) tolerance, as seen in assays with spinach [ 95 ].
3.4. Glycine Betaine (GB)
GB (N,N′,N′′-trimethylglycine) is an amphoteric quaternary ammonium compatible osmolyte that confers tolerance under environmental stress [ 96 , 97 ]. Its main beneficial effects are through osmotic adjustment and stabilisation of the photosystem II complex, amongst others [ 96 , 98 ]. Not all plants are able to accumulate GB under abiotic stress, although crops and cultivars that produce higher amounts of GB are usually more tolerant to abiotic stress [ 97 ].
Foliar application of GB to safflower ( Carthamus tinctorius ) plants subjected to salinity stress and grown under greenhouse conditions induced higher chlorophyll b and carotenoid levels, as well as a higher Fv/Fm rate. Moreover, a decrease in Na + levels in leaf tissue was observed, indicating that GB application has positive effects in the response of safflower to abiotic stress [ 99 ]. Exogenous application of GB in lettuce ( Lactuca sativa ) grown under salt stress and greenhouse conditions also resulted in higher tolerance to salinity, reflected in increased DW, organic acid, and amino acid levels when compared to control plants [ 100 ].
Application of GB to enhance abiotic stress tolerance on plants grown under field conditions has also been promising. In pea, GB application ameliorated the effects of drought stress by increasing yield and soluble protein concentration. This study evaluated short- and long-term drought stress, at different developmental stages, generating comparable scenarios to those found in field conditions [ 101 ]. In wheat, a field study using 19 genotypes grown under prolonged drought stress showed that foliar applications of this metabolite improved flag leaf net photosynthetic rate and stomatal conductance when compared to stress conditions alone. Although significant differences in chlorophylls and carotenoid accumulation were not observed, increases in AA were detected when compared to plants subjected to drought. However, responses to GB were genotype specific [ 102 ]. However, the effects of GB may be species specific—for example, no improvements were observed in growth or yield when applied to tomatoes [ 103 ], which may also be related to the dose used, the time of application, and the time of tissue collection.
Like the effect of AA on banana shelf quality [ 94 ], studies have shown that GB application also has a positive impact in the post-harvest life of commercial fruits. Immersion of hawthorn berries ( Crataegus monogyna ) in different GB concentrations and stored at 1 °C for 20 days suffered 25% less chilling injury compared to untreated controls. Application also caused significant increases in endogenous GB, AA, and proline levels, and raised activity levels of ROS scavenging enzymes (Razavi et al., 2018). Similar results were obtained in papaya ( Carica papaya ) immersed in GB after harvesting but prior to cold storage (6° for 40 days) [ 104 ].
Thus, GB not only alleviates damage produced by abiotic factors, but also enhances post-harvest life. Progress is being made in additional field studies that are identifying the species and genotypes that respond best to GB.
3.5. Alpha-Tocopherol (α-Toc)
Tocopherols are lipophilic compounds belonging to the vitamin E family. The four isomers α, β, γ, and δ play important roles in plant growth, stress responses, senescence, and signal transduction, among other processes [ 105 ]. Of these compounds, α-tocopherol (α-Toc) is the most abundant and is present in chloroplasts. One of its main functions is to prevent oxidative damage to thylakoid membranes by quenching and scavenging singlet oxygen (one α-Toc quenches approximately 120 singlet oxygen molecules) [ 106 ] and scavenging lipid peroxyl radicals, maintaining photosynthetic membrane integrity.
In recent years, the role of α-Toc in stress tolerance in crops has been examined. Most studies have investigated the beneficial effects of foliar α-Toc application during saline or drought stress. In a field experiment, Semida et al. reported a significant increase in growth parameters such as shoot length, number of leaves, leaf area, shoot DW, and FW in onion (considered a salt-sensitive crop) grown under saline condition [ 107 ] when α-Toc was used. The time of application also influences the effects of α-Toc on plant development. Lalarukh et al. [ 108 ] treated seeds of two sunflower ( Helianthus annuus ) varieties with different α-Toc concentrations and grew them in pots under NaCl treatment. Given that sunflower is moderately tolerant to salt stress, it is a good candidate for growing in saline soils. α-Toc seed priming increased shoot and root FW and shoot and root length, although the precise effect depended on both the cultivar and the metabolite concentration. It also improved yield: the authors reported up to 33 and 16% increases in total achene weight and hundred achene weights, respectively [ 109 ]. Similar results were obtained in wheat and mungbean, both grown under natural field conditions, when this metabolite was applied to plants under a water-stress regime [ 110 , 111 ]. Regarding yield improvement, α-Toc increased the number of seeds, pods, weight of ripened pods, and weight of 100 mungbean seeds [ 111 ]. In wheat, yield per plant increased by 18% when α-Toc was applied under water-stress conditions, mirrored by improvements in nutritional quality due to rises in α-, β-, γ-tocopherols, phenolics, and flavonoids [ 110 ]. Foliar application of this secondary metabolite also raised chlorophyll a and b levels and antioxidant enzyme activities in mungbean, wheat, and onions under saline stress conditions [ 107 , 110 , 111 ].
α-Toc has thus proven to enhance growth parameters and yield in crop species grown under field conditions and drought, indicating a potential use in commercial formulations. However, its beneficial effects in crops under other kinds of abiotic stress have yet to be addressed.
3.6. Melatonin
Melatonin (N-acetyl-5-methoxytryptamine) is a tryptophan derivative that was discovered in plants in 1995 [ 112 , 113 ]. Although its role in physiological processes in mammals has been extensively described [ 114 ], its function and action mechanism in crops is still being elucidated. Melatonin participates in different developmental processes, such as seed germination, growth regulation, fruit ripening, and rhizogenesis, among others [ 115 ]. Because melatonin also presents natural antioxidant capacity and delays leaf senescence, it has been proposed as a potential biostimulant for crops. An excellent review on melatonin and its function in the regulation of the plant antioxidant machinery in stressed conditions was recently published [ 116 ], thus, this section will focus on physiological parameters.
Melatonin treatment of salt-stressed tomato plants in growth chamber conditions improved growth parameters, generated higher chlorophyll a and b accumulation, and increased the activities of carbonic anhydrase and rubisco [ 117 ]. This is consistent with previous reports in other species, where melatonin treatment downregulated the expression of chlorophyllase and pheophorbide a oxygenase, two enzymes involved in chlorophyll degradation [ 118 ], thus improving photosynthetic efficiency of plants under cold stress [ 119 ].
Moreover, melatonin application increased proline levels and the activity of the enzyme required for its biosynthesis, P5CS [ 117 ]. Crosstalk of melatonin with other phytohormones, including auxin, cytokinin, gibberellins, abscisic acid, ethylene, jasmonic acid, and salicylic acid, has been determined [ 120 ]. It is a ROS scavenger and also increases the activities of antioxidant enzymes and metabolite contents [ 119 , 121 , 122 ]. Taking advantage of this crosstalk might optimise its use as a biostimulant for crops cultivated under abiotic stress. Melon seedlings ( Cucumis melo ) subjected to cold stress in growth chamber conditions presented higher chlorophyll a and b contents, net photosynthetic rate, stomatal conductance, and transpiration rate when treated with melatonin, with an optimum dose of 200 µM [ 123 ].
Other benefits attractive to the agronomical industry may be associated with abiotic stress tolerance. For instance, table grape bunches ( Vitis vinifera ) treated with melatonin presented a significantly lower berry abscission rate and a reduced rotten index, as well as more extended post-harvest life [ 124 ].
Whether melatonin should be considered a new phytohormone or a metabolite is currently under discussion [ 115 ]. Nevertheless, field trials evaluating the effects of melatonin on crops subjected to abiotic stress should be carried out in order to assess its potential impact on the yield of other species.
4. Conclusions and Future Prospects
Application of primary and secondary metabolites naturally found in plants has proven to be effective at ameliorating abiotic stress damage in crops. A variety of effects are observed in applying chemically diverse natural compounds. Some effects are common to most crop species studied, whereas others are species and even genotype specific (such as GB application in wheat [ 102 ] and Thin, 2015). Moreover, certain metabolites might perform better at alleviating specific types of stress: GSH seems to induce more tolerance to heavy metal stress, especially Hg [ 52 ]. Some of the differences observed within application of the same metabolite can be attributed to experimental design, the duration of stress, and the stage of plant development at the time of treatment. However, as Paracelsus noted, “the dose makes the poison”, and plant metabolites are not the exception (Paracelsus, “The Third Defense”). Careful consideration must be given to the dosage used, as excess of a certain metabolite might have detrimental effects on plant growth and yield.
An interesting development is the application of combinations of plant metabolites. For example, the use of ascorbic acid (AA) in addition to citric acid (CA) (2:1) had a particularly beneficial effect on stressed plants. In salt-stressed cowpea ( Vigna sinensis ; 75 mM) foliar application of AA + CA increased parameters FW and DW by 120 and 131%, respectively, as well as the number of pods and yield per plant, compared with the control (salt-stress alone). Improvements in photosynthetic pigment levels and the content of nitrogen/phosphorus/potassium were also observed, leading to greater tolerance to salt stress [ 125 ]. A similar effect occurred in apple ( Malus x domestica cv. Red Spur); AA + CA treatment increased total anthocyanin content, the total antioxidant capacity, and the activity of antioxidant enzymes (CAT), and decreased activity of polyphenol oxidase (responsible for flesh oxidation), resulting in improved nutritional quality and fruit colour [ 126 ]. These studies show that the combination of two natural metabolites (in these cases, AA + CA) is beneficial for plant development and enhancing the commercial characteristics of crops.
Although the use of natural primary and secondary metabolites has proven useful to improve abiotic stress tolerance in crops, there is a limitation to their chemical diversity. To further broaden the spectrum of compounds that can be used for this purpose, chemical genomics can be used. As the discovery of new compounds to improve abiotic stress tolerance in crops is needed, this technique has been applied to the challenge of identifying novel molecules that promote plant growth under stress conditions. Chemical genomics allows the screening of thousands of compounds from chemical libraries, based on leaf and root growth parameters in a cost- and time-efficient manner [ 127 ] [ 128 ]. This approach has proven to be effective for evaluating and selecting new compounds that could modulate plant growth under stress conditions in crops [ 127 ], and the further exploration of chemical genomics will no doubt be invaluable for progress in this field.
For several of the metabolites presented, the scalability and cost of application in crops cultivated in the field must still be evaluated. Nevertheless, in general, they are readily available for widespread implementation, and a concentrated formula could be applied in the field with the application equipment that producers already use. Although field experiments are being carried out to test the effectiveness of plant metabolites on yield, there is still the need to evaluate their potential interactions with other chemicals (both natural and artificially added) in irrigation waters. Furthermore, the availability and production costs should be built into the equation to determine their viability as commercial products. Finally, there are salt-susceptible crop varieties that present unique or customer-valued attributes, and for these reasons, they cannot be replaced, or it is difficult to do so. In such cases, the application of different metabolites (alone or in combination with others) will allow its cultivation in saline soil conditions, maintaining its yield.
Acknowledgments
We thank all members of the Centro de Biología Molecular Vegetal for helpful and constructive discussions.
Author Contributions
Conceptualisation, F.G. and M.H.; writing—original draft preparation, F.G. and K.O.-H.; writing—review and editing, F.G., K.O.-H., C.S. and M.H.; funding acquisition, M.H. and C.S. All authors have read and agreed to the published version of the manuscript.
This research was funded by CONICYT PIA ACT192073 (MH, CS) and Fondecyt 1181198 (MH).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
- 1. Rockström J., Williams J., Daily G., Noble A., Matthews N., Gordon L., Wetterstrand H., DeClerck F., Shah M., Steduto P., et al. Sustainable intensification of agriculture for human prosperity and global sustainability. Ambio. 2017;46:4–17. doi: 10.1007/s13280-016-0793-6. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 2. Döös B.R. Population growth and loss of arable land. Glob. Environ. Chang. 2002;12:303–311. doi: 10.1016/S0959-3780(02)00043-2. [ DOI ] [ Google Scholar ]
- 3. Hofmann D.J., Butler J.H., Tans P.P. A new look at atmospheric carbon dioxide. Atmos. Environ. 2009;43:2084–2086. doi: 10.1016/j.atmosenv.2008.12.028. [ DOI ] [ Google Scholar ]
- 4. U.S. Global Change Research Program . Global Climate Change Impacts in the United States: A State of Knowledge Report. Cambridge University Press; Cambridge, UK; New York, NY, USA: 2009. [ Google Scholar ]
- 5. Bot A.J., Nachtergaele F.O., Young A. Land Resource Potential and Constraints at Regional and Country Levels. World Soil Resources Report; FAO; Rome, Italy: 2000. p. 122. [ Google Scholar ]
- 6. Thornton P.K., Ericksen P.J., Herrero M., Challinor A.J. Climate variability and vulnerability to climate change: A review. Glob. Chang. Biol. 2014;20:3313–3328. doi: 10.1111/gcb.12581. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 7. Ahmad B. Effects of Salinity on Yield and Component Characters in Canola (Brassica napus L.) Cultivars. Not. Sci. Biol. 2010;2:81–83. doi: 10.15835/nsb.2.1.3560. [ DOI ] [ Google Scholar ]
- 8. Panta S., Flowers T., Lane P., Doyle R., Haros G., Shabala S. Halophyte agriculture: Success stories. Environ. Exp. Bot. 2014;107:71–83. doi: 10.1016/j.envexpbot.2014.05.006. [ DOI ] [ Google Scholar ]
- 9. Manzoor J., Sharma M., Wani K.A. Heavy metals in vegetables and their impact on the nutrient quality of vegetables: A review. J. Plant Nutr. 2018;41:1744–1763. doi: 10.1080/01904167.2018.1462382. [ DOI ] [ Google Scholar ]
- 10. FAO . Status of the World’s Soil Resources: Main Report. FAO, ITPS; Rome, Italy: 2015. [ Google Scholar ]
- 11. Nadarajah K.K. ROS homeostasis in abiotic stress tolerance in plants. Int. J. Mol. Sci. 2020;21:5208. doi: 10.3390/ijms21155208. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 12. You J., Chan Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015;6:1092. doi: 10.3389/fpls.2015.01092. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 13. Apel K., Hirt H. REACTIVE OXYGEN SPECIES: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004;55:373–399. doi: 10.1146/annurev.arplant.55.031903.141701. [ DOI ] [ PubMed ] [ Google Scholar ]
- 14. Foyer C.H., Noctor G. Redox regulation in photosynthetic organisms: Signaling, acclimation, and practical implications. Antioxid. Redox Signal. 2009;11:861–905. doi: 10.1089/ars.2008.2177. [ DOI ] [ PubMed ] [ Google Scholar ]
- 15. Regulation (EU) 2019/1009 of the European Parliament and of the Council of 5 June 2019 Laying Down Rules on the Making Available on the Market of EU Fertilising Products and Amending Regulations (EC) No 1069/2009 and (EC) No 1107/2009 and Repealing Regulation (EC) No 2003/2003 (Text with EEA Relevance) Vol. 170. European Parliament, Council of the European Union; Luxembourg: 2019. pp. 1–114. [ Google Scholar ]
- 16. Kumar K., Gambhir G., Dass A., Tripathi A.K., Singh A., Jha A.K., Yadava P., Choudhary M., Rakshit S. Genetically modified crops: Current status and future prospects. Planta. 2020;251:91. doi: 10.1007/s00425-020-03372-8. [ DOI ] [ PubMed ] [ Google Scholar ]
- 17. McDougall P. The Cost and Time Involved in the Discovery, Development and Authorisation of a New Plant Biotechnology Derived Trait. Consultancy Study for Crop Life International by P McDougall; Pathhead, Midlothian: 2011. pp. 1–24. [ Google Scholar ]
- 18. Waltz E. Gene-edited CRISPR mushroom escapes US regulation. Nat. News. 2016;532:293. doi: 10.1038/nature.2016.19754. [ DOI ] [ PubMed ] [ Google Scholar ]
- 19. Jordan N.R., Dorn K.M., Smith T.M., Wolf K.E., Ewing P.M., Fernandez A.L., Runck B.C., Williams A., Lu Y., Kuzma J. A cooperative governance network for crop genome editing. EMBO Rep. 2017;18:1683–1687. doi: 10.15252/embr.201744394. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 20. Ahmar S., Gill R.A., Jung K.-H., Faheem A., Qasim M.U., Mubeen M., Zhou W. Conventional and Molecular Techniques from Simple Breeding to Speed Breeding in Crop Plants: Recent Advances and Future Outlook. Int. J. Mol. Sci. 2020;21:2590. doi: 10.3390/ijms21072590. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 21. Aremu A.O., Fawole O.A., Makunga N.P., Masondo N.A., Moyo M., Buthelezi N.M.D., Amoo S.O., Spíchal L., Doležal K. Applications of Cytokinins in Horticultural Fruit Crops: Trends and Future Prospects. Biomolecules. 2020;10:1222. doi: 10.3390/biom10091222. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 22. Javid M.G., Sorooshzadeh A., Moradi F., Modarres Sanavy S.A.M., Allahdadi I. The Role of Phytohormones in Alleviating Salt Stress in Crop Plants. Aust. J. Crop Sci. 2011;5:726. [ Google Scholar ]
- 23. Ribba T., Garrido-Vargas F., O’Brien J.A. Auxin-mediated responses under salt stress: From developmental regulation to biotechnological applications. J. Exp. Bot. 2020;71:3843–3853. doi: 10.1093/jxb/eraa241. [ DOI ] [ PubMed ] [ Google Scholar ]
- 24. Sah S.K., Reddy K.R., Li J. Abscisic Acid and Abiotic Stress Tolerance in Crop Plants. Front. Plant Sci. 2016;7:571. doi: 10.3389/fpls.2016.00571. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 25. Wani S.H., Kumar V., Shriram V., Sah S.K. Phytohormones and their metabolic engineering for abiotic stress tolerance in crop plants. Crop J. 2016;4:162–176. doi: 10.1016/j.cj.2016.01.010. [ DOI ] [ Google Scholar ]
- 26. Geilfus C.-M. Controlled Environment Horticulture: Improving Quality of Vegetables and Medicinal Plants. Springer International Publishing; Cham, Switzerland: 2019. [ Google Scholar ]
- 27. Wink M. eLS. John Wiley & Sons, Ltd.; Chichester, UK: 2016. Evolution of Secondary Plant Metabolism; pp. 1–11. [ Google Scholar ]
- 28. Nabavi S.M., Šamec D., Tomczyk M., Milella L., Russo D., Habtemariam S., Suntar I., Rastrelli L., Daglia M., Xiao J., et al. Flavonoid biosynthetic pathways in plants: Versatile targets for metabolic engineering. Biotechnol. Adv. 2020;38:107316. doi: 10.1016/j.biotechadv.2018.11.005. [ DOI ] [ PubMed ] [ Google Scholar ]
- 29. Szabados L., Savouré A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010;15:89–97. doi: 10.1016/j.tplants.2009.11.009. [ DOI ] [ PubMed ] [ Google Scholar ]
- 30. Mansour M.M.F., Ali E.F. Evaluation of proline functions in saline conditions. Phytochemistry. 2017;140:52–68. doi: 10.1016/j.phytochem.2017.04.016. [ DOI ] [ PubMed ] [ Google Scholar ]
- 31. Teh C.-Y., Shaharuddin N.A., Ho C.-L., Mahmood M. Exogenous proline significantly affects the plant growth and nitrogen assimilation enzymes activities in rice (Oryza sativa) under salt stress. Acta Physiol. Plant. 2016;38:151. doi: 10.1007/s11738-016-2163-1. [ DOI ] [ Google Scholar ]
- 32. Tabssum F., Zaman Q., Chen Y., Riaz U., Ashraf W., Aslam A., Ehsan N., Nawaz R., Aziz H., Shah S. Exogenous Application of Proline Improved Salt Tolerance in Rice through Modulation of Antioxidant Activities. Pak. J. Agric. Res. 2019;32:140–151. doi: 10.17582/journal.pjar/2019/32.1.140.151. [ DOI ] [ Google Scholar ]
- 33. Dawood M.G., Taie H.A.A., Nassar R.M.A., Abdelhamid M.T., Schmidhalter U. The changes induced in the physiological, biochemical and anatomical characteristics of Vicia faba by the exogenous application of proline under seawater stress. South Afr. J. Bot. 2014;93:54–63. doi: 10.1016/j.sajb.2014.03.002. [ DOI ] [ Google Scholar ]
- 34. Rady M.M., Taha R.S., Mahdi A.H.A. Proline enhances growth, productivity and anatomy of two varieties of Lupinus termis L. grown under salt stress. South Afr. J. Bot. 2016;102:221–227. doi: 10.1016/j.sajb.2015.07.007. [ DOI ] [ Google Scholar ]
- 35. Saradhi P.P., Mohanty P. Involvement of proline in protecting thylakoid membranes against free radical-induced photodamage. J. Photochem. Photobiol. B. 1997;38:253–257. doi: 10.1016/S1011-1344(96)07470-2. [ DOI ] [ Google Scholar ]
- 36. Singh M., Pratap Singh V., Dubey G., Mohan Prasad S. Exogenous proline application ameliorates toxic effects of arsenate in Solanum melongena L. seedlings. Ecotoxicol. Environ. Saf. 2015;117:164–173. doi: 10.1016/j.ecoenv.2015.03.021. [ DOI ] [ PubMed ] [ Google Scholar ]
- 37. Yu X.-Z., Lin Y.-J., Fan W.-J., Lu M.-R. The role of exogenous proline in amelioration of lipid peroxidation in rice seedlings exposed to Cr(VI) Int. Biodeterior. Biodegrad. 2017;123:106–112. doi: 10.1016/j.ibiod.2017.06.010. [ DOI ] [ Google Scholar ]
- 38. Noreen S., Akhter M.S., Yaamin T., Arfan M. The ameliorative effects of exogenously applied proline on physiological and biochemical parameters of wheat (Triticum aestivum L.) crop under copper stress condition. J. Plant Interact. 2018;13:221–230. doi: 10.1080/17429145.2018.1437480. [ DOI ] [ Google Scholar ]
- 39. Mustafa A., Imran M., Ashraf M., Mahmood K. Perspectives of using L-Tryptophan for improving productivity of agricultural crops: A Review. Pedosphere. 2018;28:16–34. doi: 10.1016/S1002-0160(18)60002-5. [ DOI ] [ Google Scholar ]
- 40. Jamil M., Kharal M.A., Ahmad M., Abbasi G.H., Nazli F., Hussain A., Akhtar M.F.-Z. Inducing salinity tolerance in red pepper (Capsicum annuum L.) through exogenous application of proline and L-tryptophan. Soil Environ. 2018;37:160–168. [ Google Scholar ]
- 41. Hozayn M., Abd-Elmonem A.A., Samaha G.M. The physiological effect of pre-soaking with tryptophan on sugar beet (Beta vulgaris L.) productivity under different levels of salinity stresses. Bull. Natl. Res. Cent. 2020;44:65. doi: 10.1186/s42269-020-00324-w. [ DOI ] [ Google Scholar ]
- 42. Jamil M., Ahamd M., Anwar F., Zahir Z., Ali Kharal M., Nazli F. Inducing drought tolerance in wheat through combined use of L-tryptophan and pseudomonas fluorescens. Pak. J. Agric. Sci. 2018;55:331–337. doi: 10.21162/PAKJAS/18.4980. [ DOI ] [ Google Scholar ]
- 43. Farooq H., Asghar H.N., Khan M.Y., Saleem M., Zahir Z.A. Auxin-mediated growth of rice in cadmium-contaminated soil. Turk. J. Agric. For. 2015;39:272–276. doi: 10.3906/tar-1405-54. [ DOI ] [ Google Scholar ]
- 44. Hanci F., Çingi M., Akinci H. Influence of L-Tryptophan and Melatonin on Germination of Onion and Leek Seeds at Different Temperatures. Türkiye Tarımsal Araştırmalar Derg. 2019;6:214–221. doi: 10.19159/tutad.559617. [ DOI ] [ Google Scholar ]
- 45. Noctor G., Mhamdi A., Chaouch S., Han Y., Neukermans J., Marquez-Garcia B., Queval G., Foyer C.H. Glutathione in plants: An integrated overview. Plant Cell Environ. 2012;35:454–484. doi: 10.1111/j.1365-3040.2011.02400.x. [ DOI ] [ PubMed ] [ Google Scholar ]
- 46. Hasanuzzaman M., Nahar K., Anee T.I., Fujita M. Glutathione in plants: Biosynthesis and physiological role in environmental stress tolerance. Physiol. Mol. Biol. Plants. 2017;23:249–268. doi: 10.1007/s12298-017-0422-2. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 47. Hasanuzzaman M., Nahar K., Rahman A., Mahmud J.A., Alharby H.F., Fujita M. Exogenous glutathione attenuates lead-induced oxidative stress in wheat by improving antioxidant defense and physiological mechanisms. J. Plant Interact. 2018;13:203–212. doi: 10.1080/17429145.2018.1458913. [ DOI ] [ Google Scholar ]
- 48. Nahar K., Hasanuzzaman M., Alam M.M., Fujita M. Roles of exogenous glutathione in antioxidant defense system and methylglyoxal detoxification during salt stress in mung bean. Biol. Plant. 2015;59:745–756. doi: 10.1007/s10535-015-0542-x. [ DOI ] [ Google Scholar ]
- 49. Zhou Y., Wen Z., Zhang J., Chen X., Cui J., Xu W., Liu H. Exogenous glutathione alleviates salt-induced oxidative stress in tomato seedlings by regulating glutathione metabolism, redox status, and the antioxidant system. Sci. Hortic. 2017;220:90–101. doi: 10.1016/j.scienta.2017.02.021. [ DOI ] [ Google Scholar ]
- 50. Akram S., Siddiqui M.N., Hussain B.M.N., Al Bari M.A., Mostofa M.G., Hossain M.A., Tran L.-S.P. Exogenous glutathione modulates salinity tolerance of soybean [Glycine max (L.) Merrill] at reproductive stage. J. Plant Growth Regul. 2017;36:877–888. doi: 10.1007/s00344-017-9691-9. [ DOI ] [ Google Scholar ]
- 51. Khan M., Daud M.K., Basharat A., Khan M.J., Azizullah A., Muhammad N., Muhammad N., ur Rehman Z., Zhu S.J. Alleviation of lead-induced physiological, metabolic, and ultramorphological changes in leaves of upland cotton through glutathione. Environ. Sci. Pollut. Res. 2016;23:8431–8440. doi: 10.1007/s11356-015-5959-4. [ DOI ] [ PubMed ] [ Google Scholar ]
- 52. Kim Y.-O., Bae H.-J., Cho E., Kang H. Exogenous Glutathione Enhances Mercury Tolerance by Inhibiting Mercury Entry into Plant Cells. Front. Plant Sci. 2017;8:683. doi: 10.3389/fpls.2017.00683. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 53. Nahar K., Hasanuzzaman M., Alam M.M., Fujita M. Exogenous glutathione confers high temperature stress tolerance in mung bean (Vigna radiata L.) by modulating antioxidant defense and methylglyoxal detoxification system. Environ. Exp. Bot. 2015;112:44–54. doi: 10.1016/j.envexpbot.2014.12.001. [ DOI ] [ Google Scholar ]
- 54. Kumar P., Dubey K.K. New and Future Developments in Microbial Biotechnology and Bioengineering. Elsevier; Amsterdam, The Netherlands: 2019. Citric acid cycle regulation: Back bone for secondary metabolite production; pp. 165–181. [ Google Scholar ]
- 55. Soccol C.R., Vandenberghe L.P.S., Rodrigues C., Pandey A. New perspectives for citric acid production and application. Food Technol. Biotechnol. 2006;44:141–149. [ Google Scholar ]
- 56. Pérez-Labrada F., Benavides-Mendoza A., Valdez-Aguilar L.A., Robledo-Torres V. Citric acid in the nutrient solution increases the mineral absorption in potted tomato grown in calcareous soil. Pak. J. Bot. 2016;48:67–74. [ Google Scholar ]
- 57. El-Hawary M.M., Nashed M.E. Effect of Foliar Application by some Antioxidants on Growth and Productivity of Maize under Saline Soil Conditions. J. Plant Prod. 2019;10:93–99. doi: 10.21608/jpp.2019.36238. [ DOI ] [ Google Scholar ]
- 58. Faraz A., Faizan M., Sami F., Siddiqui H., Hayat S. Supplementation of Salicylic Acid and Citric Acid for Alleviation of Cadmium Toxicity to Brassica juncea. J. Plant Growth Regul. 2020;39:641–655. doi: 10.1007/s00344-019-10007-0. [ DOI ] [ Google Scholar ]
- 59. Mallhi Z.I., Rizwan M., Mansha A., Ali Q., Asim S., Ali S., Hussain A., Alrokayan S.H., Khan H.A., Alam P., et al. Citric acid enhances plant growth, photosynthesis, and phytoextraction of lead by alleviating the oxidative stress in Castor beans. Plants. 2019;8:525. doi: 10.3390/plants8110525. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 60. Zaheer I.E., Ali S., Rizwan M., Farid M., Shakoor M.B., Gill R.A., Najeeb U., Iqbal N., Ahmad R. Citric acid assisted phytoremediation of copper by Brassica napus L. Ecotoxicol. Environ. Saf. 2015;120:310–317. doi: 10.1016/j.ecoenv.2015.06.020. [ DOI ] [ PubMed ] [ Google Scholar ]
- 61. Ben Massoud M., Sakouhi L., Chaoui A. Effect of plant growth regulators, calcium and citric acid on copper toxicity in pea seedlings. J. Plant Nutr. 2019;42:1230–1242. doi: 10.1080/01904167.2019.1609506. [ DOI ] [ Google Scholar ]
- 62. Parveen A., Saleem M.H., Kamran M., Haider M.Z., Chen J.-T., Malik Z., Rana M.S., Hassan A., Hur G., Javed M.T., et al. Effect of citric acid on growth, ecophysiology, chloroplast ultrastructure, and phytoremediation potential of jute (Corchorus capsularis L.) seedlings exposed to copper stress. Biomolecules. 2020;10:592. doi: 10.3390/biom10040592. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 63. Khatun M.R., Mukta R.H., Islam M.A., Huda A.K.M.N. Insight into citric acid-induced chromium detoxification in rice (Oryza sativa L.) Int. J. Phytoremediation. 2019;21:1234–1240. doi: 10.1080/15226514.2019.1619162. [ DOI ] [ PubMed ] [ Google Scholar ]
- 64. Maqbool A., Ali S., Rizwan M., Ishaque W., Rasool N., Rehman M.Z.u., Bashir A., Abid M., Wu L. Management of tannery wastewater for improving growth attributes and reducing chromium uptake in spinach through citric acid application. Environ. Sci. Pollut. Res. 2018;25:10848–10856. doi: 10.1007/s11356-018-1352-4. [ DOI ] [ PubMed ] [ Google Scholar ]
- 65. Munnik T., Vermeer J.E.M. Osmotic stress-induced phosphoinositide and inositol phosphate signalling in plants. Plant Cell Environ. 2010;33:655–669. doi: 10.1111/j.1365-3040.2009.02097.x. [ DOI ] [ PubMed ] [ Google Scholar ]
- 66. Chatterjee J., Majumder A.L. Salt-induced abnormalities on root tip mitotic cells of Allium cepa: Prevention by inositol pretreatment. Protoplasma. 2010;245:165–172. doi: 10.1007/s00709-010-0170-4. [ DOI ] [ PubMed ] [ Google Scholar ]
- 67. Gerke J. Phytate (Inositol Hexakisphosphate) in soil and phosphate acquisition from inositol phosphates by higher plants. A Review. Plants. 2015;4:253–266. doi: 10.3390/plants4020253. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 68. Kaya C., Sonmez O., Aydemir S., Ashraf M., Dikilitas M. Exogenous application of mannitol and thiourea regulates plant growth and oxidative stress responses in salt-stressed maize (Zea mays L.) J. Plant Interact. 2013;8:234–241. doi: 10.1080/17429145.2012.725480. [ DOI ] [ Google Scholar ]
- 69. Ramadan M.E., Shalaby O.A. Effect of salicylic acid and mannitol on white cabbage plants under saline conditions. J. Plant Prod. 2018;9:397–402. doi: 10.21608/jpp.2018.35766. [ DOI ] [ Google Scholar ]
- 70. Habiba U., Ali S., Rizwan M., Ibrahim M., Hussain A., Shahid M.R., Alamri S.A., Alyemeni M.N., Ahmad P. Alleviative role of exogenously applied mannitol in maize cultivars differing in chromium stress tolerance. Environ. Sci. Pollut. Res. 2019;26:5111–5121. doi: 10.1007/s11356-018-3970-2. [ DOI ] [ PubMed ] [ Google Scholar ]
- 71. Adrees M., Ali S., Iqbal M., Aslam Bharwana S., Siddiqi Z., Farid M., Ali Q., Saeed R., Rizwan M. Mannitol alleviates chromium toxicity in wheat plants in relation to growth, yield, stimulation of anti-oxidative enzymes, oxidative stress and Cr uptake in sand and soil media. Ecotoxicol. Environ. Saf. 2015;122:1–8. doi: 10.1016/j.ecoenv.2015.07.003. [ DOI ] [ PubMed ] [ Google Scholar ]
- 72. Landi M., Margaritopoulou T., Papadakis I.E., Araniti F. Boron toxicity in higher plants: An update. Planta. 2019;250:1011–1032. doi: 10.1007/s00425-019-03220-4. [ DOI ] [ PubMed ] [ Google Scholar ]
- 73. Coskun Y., Olgunsoy P., Karatas N., Bulut F., Yarar F. Mannitol application alleviates boron toxicity in wheat seedlings. Commun. Soil Sci. Plant Anal. 2014;45:944–952. doi: 10.1080/00103624.2013.867054. [ DOI ] [ Google Scholar ]
- 74. Issa D.B., Alturki S.M., Sajyan T.K., Sassine Y.N. Sorbitol and lithovit-guano25 mitigates the adverse effects of salinity on eggplant grown in pot experiment. Agron. Res. 2020;18:113–126. doi: 10.15159/AR.20.075. [ DOI ] [ Google Scholar ]
- 75. Theerakulpisut P., Gunnula W. Exogenous sorbitol and trehalose mitigated salt stress damage in Salt- sensitive but not Salt-tolerant rice seedlings. Asian J. Crop Sci. 2012;4:165–170. doi: 10.3923/ajcs.2012.165.170. [ DOI ] [ Google Scholar ]
- 76. Kokitkar S.S., Chavan L.R., Bhagat S.A. Alleviation of adverse effects of salt stress in triticum aestivum by foliar application of compatible solutes. IJRBAT. 2017;V:272–276. [ Google Scholar ]
- 77. Hu L., Zhou K., Li Y., Chen X., Liu B., Li C., Gong X., Ma F. Exogenous myo-inositol alleviates salinity-induced stress in Malus hupehensis Rehd. Plant Physiol. Biochem. 2018;133:116–126. doi: 10.1016/j.plaphy.2018.10.037. [ DOI ] [ PubMed ] [ Google Scholar ]
- 78. Yildizli A., Çevik S., Ünyayar S. Effects of exogenous myo-inositol on leaf water status and oxidative stress of Capsicum annuum under drought stress. Acta Physiol. Plant. 2018;40:122. doi: 10.1007/s11738-018-2690-z. [ DOI ] [ Google Scholar ]
- 79. Araya-Flores J., Miranda S., Covarrubias M.P., Stange C., Handford M. Solanum lycopersicum (tomato) possesses mitochondrial and plastidial lipoyl synthases capable of increasing lipoylation levels when expressed in bacteria. Plant Physiol. Biochem. 2020;151:264–270. doi: 10.1016/j.plaphy.2020.03.031. [ DOI ] [ PubMed ] [ Google Scholar ]
- 80. Yıldız M., Akçalı N., Terzi H. Proteomic and biochemical responses of canola (Brassica napus L.) exposed to salinity stress and exogenous lipoic acid. J. Plant Physiol. 2015;179:90–99. doi: 10.1016/j.jplph.2015.03.006. [ DOI ] [ PubMed ] [ Google Scholar ]
- 81. Elrys A.S., Abdo A.I.E., Abdel-Hamed E.M.W., Desoky E.-S.M. Integrative application of licorice root extract or lipoic acid with fulvic acid improves wheat production and defenses under salt stress conditions. Ecotoxicol. Environ. Saf. 2020;190:110144. doi: 10.1016/j.ecoenv.2019.110144. [ DOI ] [ PubMed ] [ Google Scholar ]
- 82. Sezgin A., Altuntaş C., Demiralay M., Cinemre S., Terzi R. Exogenous alpha lipoic acid can stimulate photosystem II activity and the gene expressions of carbon fixation and chlorophyll metabolism enzymes in maize seedlings under drought. J. Plant Physiol. 2019;232:65–73. doi: 10.1016/j.jplph.2018.11.026. [ DOI ] [ PubMed ] [ Google Scholar ]
- 83. Turk H., Erdal S., Karayel U., Dumlupinar R. Attenuation of lead toxicity by promotion of tolerance mechanism in wheat roots by lipoic acid. Cereal Res. Commun. 2018;46:424–435. doi: 10.1556/0806.46.2018.020. [ DOI ] [ Google Scholar ]
- 84. Wang J., Zhang Z., Huang R. Regulation of ascorbic acid synthesis in plants. Plant Signal. Behav. 2013;8:e24536. doi: 10.4161/psb.24536. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 85. Hossain M.A., Munné-Bosch S., Burritt D.J., Diaz-Vivancos P., Fujita M., Lorence A. Springer International Publishing; Cham, Switzerland: 2017. Ascorbic Acid in Plant Growth, Development and Stress Tolerance. [ Google Scholar ]
- 86. Bilska K., Wojciechowska N., Alipour S., Kalemba E.M. Ascorbic Acid—The Little-Known Antioxidant in Woody Plants. Antioxidants. 2019;8:645. doi: 10.3390/antiox8120645. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 87. Aziz A., Akram N.A., Ashraf M. Influence of natural and synthetic vitamin C (Ascorbic acid) on primary and secondary metabolites and associated metabolism in quinoa (Chenopodium quinoa Willd.) plants under water deficit regimes. Plant Physiol. Biochem. 2018;123:192–203. doi: 10.1016/j.plaphy.2017.12.004. [ DOI ] [ PubMed ] [ Google Scholar ]
- 88. Penella C., Calatayud Á., Melgar J.C. Ascorbic acid alleviates water stress in young peach trees and improves their performance after rewatering. Front. Plant Sci. 2017;8:1627. doi: 10.3389/fpls.2017.01627. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 89. Hafez E.M., Gharib H.S. Effect of exogenous application of ascorbic acid on physiological and biochemical characteristics of wheat under water stress. Int. J. Plant Prod. 2016;10:579–596. [ Google Scholar ]
- 90. Alamri S.A., Siddiqui M.H., Al-Khaishany M.Y., Nasir Khan M., Ali H.M., Alaraidh I.A., Alsahli A.A., Al-Rabiah H., Mateen M. Ascorbic acid improves the tolerance of wheat plants to lead toxicity. J. Plant Interact. 2018;13:409–419. doi: 10.1080/17429145.2018.1491067. [ DOI ] [ Google Scholar ]
- 91. Sarwar M., Saleem M.F., Ullah N., Rizwan M., Ali S., Shahid M.R., Alamri S.A., Alyemeni M.N., Ahmad P. Exogenously applied growth regulators protect the cotton crop from heat-induced injury by modulating plant defense mechanism. Sci. Rep. 2018;8:17086. doi: 10.1038/s41598-018-35420-5. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 92. Kamal M.A., Saleem M.F., Shahid M., Awais M., Khan H.Z., Ahmed K. Ascorbic acid triggered physiochemical transformations at different phenological stages of heat-stressed Bt cotton. J. Agron. Crop Sci. 2017;203:323–331. doi: 10.1111/jac.12211. [ DOI ] [ Google Scholar ]
- 93. Elkelish A., Qari S.H., Mazrou Y.S.A., Abdelaal K.A.A., Hafez Y.M., Abu-Elsaoud A.M., Batiha G.E.-S., El-Esawi M.A., El Nahhas N. Exogenous Ascorbic acid induced chilling tolerance in tomato plants through modulating metabolism, osmolytes, antioxidants, and transcriptional regulation of catalase and heat shock proteins. Plants. 2020;9:431. doi: 10.3390/plants9040431. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 94. Lo’ay A.A., EL-Khateeb A.Y. Antioxidant enzyme activities and exogenous ascorbic acid treatment of ‘Williams’ banana during long-term cold storage stress. Sci. Hortic. 2018;234:210–219. doi: 10.1016/j.scienta.2018.02.038. [ DOI ] [ Google Scholar ]
- 95. Min K., Chen K., Arora R. A metabolomics study of ascorbic acid-induced in spinach (Spinacia oleracea L. ) Plant Direct. 2020;4:1–13. doi: 10.1002/pld3.202. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 96. Giri J. Glycinebetaine and abiotic stress tolerance in plants. Plant Signal. Behav. 2011;6:1746–1751. doi: 10.4161/psb.6.11.17801. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 97. Kurepin L.V., Ivanov A.G., Zaman M., Pharis R.P., Hurry V., Hüner N.P.A. Interaction of Glycine Betaine and Plant Hormones: Protection of the Photosynthetic Apparatus during Abiotic Stress. In: Hou H.J.M., Najafpour M.M., Moore G.F., Allakhverdiev S.I., editors. Photosynthesis: Structures, Mechanisms, and Applications. Springer International Publishing; Cham, Switzerland: 2017. pp. 185–202. [ Google Scholar ]
- 98. Huang S., Zuo T., Ni W. Important roles of glycinebetaine in stabilizing the structure and function of the photosystem II complex under abiotic stresses. Planta. 2020;251:36. doi: 10.1007/s00425-019-03330-z. [ DOI ] [ PubMed ] [ Google Scholar ]
- 99. Alasvandyari F., Mahdavi B. Effect of glycine betaine and salinity on photosynthetic pigments and ion concentration of safflower. Desert. 2018;23:265–271. [ Google Scholar ]
- 100. Shams M., Yildirim E., Ekinci M., Turan M., Dursun A., Parlakova F., Kul R. Exogenously applied glycine betaine regulates some chemical characteristics and antioxidative defence systems in lettuce under salt stress. Hortic. Environ. Biotechnol. 2016;57:225–231. doi: 10.1007/s13580-016-0021-0. [ DOI ] [ Google Scholar ]
- 101. Osman H.S. Enhancing antioxidant–yield relationship of pea plant under drought at different growth stages by exogenously applied glycine betaine and proline. Ann. Agric. Sci. 2015;60:389–402. doi: 10.1016/j.aoas.2015.10.004. [ DOI ] [ Google Scholar ]
- 102. Gupta N., Thind S. Improving photosynthetic performance of bread wheat under field drought stress by foliar applied glycine betaine. J. Agric. Sci. Tech. 2015;17:75–86. [ Google Scholar ]
- 103. Sajyan T.K., Allaw W., Shaban N., Sassine Y.N. Effect of exogenous application of glycine betaine on tomato plants subjected to salt stress. Acta Hortic. 2019;1253:41–48. doi: 10.17660/ActaHortic.2019.1253.6. [ DOI ] [ Google Scholar ]
- 104. Pan Y., Zhang S., Yuan M., Song H., Wang T., Zhang W., Zhang Z. Effect of glycine betaine on chilling injury in relation to energy metabolism in papaya fruit during cold storage. Food Sci. Nutr. 2019;7:1123–1130. doi: 10.1002/fsn3.957. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 105. Munné-Bosch S., Alegre L. The function of tocopherols and tocotrienols in plants. Crit. Rev. Plant Sci. 2002;21:31–57. doi: 10.1080/0735-260291044179. [ DOI ] [ Google Scholar ]
- 106. Gill S.S., Tuteja N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010;48:909–930. doi: 10.1016/j.plaphy.2010.08.016. [ DOI ] [ PubMed ] [ Google Scholar ]
- 107. Semida W.M., Abd El-Mageed T.A., Howladar S.M., Rady M.M. Foliar-applied alpha-tocopherol enhances salt-tolerance in onion plants by improving antioxidant defense system. Aust. J. Crop Sci. 2016;10:1030. doi: 10.21475/ajcs.2016.10.07.p7712. [ DOI ] [ Google Scholar ]
- 108. Lalarukh I. Alpha-tocopherol induced modulations in morpho-physiological attributes of sunflower (Helianthus annuus) grown under saline environment. Int. J. Agric. Biol. 2018;20:661–668. doi: 10.17957/IJAB/15.0538. [ DOI ] [ Google Scholar ]
- 109. Lalarukh I., Shahbaz M. Response of antioxidants and lipid peroxidation to exogenous application of alpha-tocopherol in sunflower (Helianthus annuus L.) under salt stress. Pak. J. Bot. 2020;52:75–83. doi: 10.30848/PJB2020-1(41). [ DOI ] [ Google Scholar ]
- 110. Ali Q., Ali S., Iqbal N., Javed M., Rizwan M., Khaliq R., Shahid S., Parveen R., Alamri S., Alyemeni M., et al. Alpha-tocopherol fertigation confers growth physio-biochemical and qualitative yield enhancement in field grown water deficit wheat (Triticum aestivum L.) Sci. Rep. 2019;9:1–15. doi: 10.1038/s41598-019-49481-7. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 111. Sadiq M., Akram N.A., Ashraf M. Impact of exogenously applied tocopherol on some key physio-biochemical and yield attributes in mungbean [Vigna radiata (L.) Wilczek] under limited irrigation regimes. Acta Physiol. Plant. 2018;40:131. doi: 10.1007/s11738-018-2711-y. [ DOI ] [ Google Scholar ]
- 112. Dubbels R., Reiter R.J., Klenke E., Goebel A., Schnakenberg E., Ehlers C., Schiwara H.W., Schloot W. Melatonin in edible plants identified by radioimmunoassay and by high performance liquid chromatography-mass spectrometry. J. Pineal Res. 1995;18:28–31. doi: 10.1111/j.1600-079X.1995.tb00136.x. [ DOI ] [ PubMed ] [ Google Scholar ]
- 113. Hattori A., Migitaka H., Iigo M., Itoh M., Yamamoto K., Ohtani-Kaneko R., Hara M., Suzuki T., Reiter R. Identification of melatonin in plants and its effects on plasma melatonin levels and binding to melatonin receptors in vertebrates. Biochem. Mol. Biol. Int. 1995;35:627–634. [ PubMed ] [ Google Scholar ]
- 114. Hardeland R., Madrid J.A., Tan D.-X., Reiter R.J. Melatonin, the circadian multioscillator system and health: The need for detailed analyses of peripheral melatonin signaling: Melatonin and peripheral oscillators. J. Pineal Res. 2012;52:139–166. doi: 10.1111/j.1600-079X.2011.00934.x. [ DOI ] [ PubMed ] [ Google Scholar ]
- 115. Arnao M.B., Hernández-Ruiz J. Melatonin: Plant growth regulator and/or biostimulator during stress? Trends Plant Sci. 2014;19:789–797. doi: 10.1016/j.tplants.2014.07.006. [ DOI ] [ PubMed ] [ Google Scholar ]
- 116. Khan A., Numan M., Khan A.L., Lee I.-J., Imran M., Asaf S., Al-Harrasi A. Melatonin: Awakening the defense mechanisms during plant oxidative stress. Plants. 2020;9:407. doi: 10.3390/plants9040407. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 117. Siddiqui M.H., Alamri S., Al-Khaishany M.Y., Khan M.N., Al-Amri A., Ali H.M., Alaraidh I.A., Alsahli A.A. Exogenous melatonin counteracts nacl-induced damage by regulating the antioxidant system, proline and carbohydrates metabolism in tomato seedlings. Int. J. Mol. Sci. 2019;20:353. doi: 10.3390/ijms20020353. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 118. Weeda S., Zhang N., Zhao X., Ndip G., Guo Y., Buck G.A., Fu C., Ren S. Arabidopsis transcriptome analysis reveals key roles of melatonin in plant defense systems. PLoS ONE. 2014;9:e93462. doi: 10.1371/journal.pone.0093462. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 119. Fan J., Hu Z., Xie Y., Chan Z., Chen K., Amombo E., Chen L., Fu J. Alleviation of cold damage to photosystem II and metabolisms by melatonin in Bermudagrass. Front. Plant Sci. 2015;6:925. doi: 10.3389/fpls.2015.00925. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 120. Arnao M.B., Hernández-Ruiz J. Melatonin: A new plant hormone and/or a plant master regulator? Trends Plant Sci. 2019;24:38–48. doi: 10.1016/j.tplants.2018.10.010. [ DOI ] [ PubMed ] [ Google Scholar ]
- 121. Manchester L.C., Coto-Montes A., Boga J.A., Andersen L.P.H., Zhou Z., Galano A., Vriend J., Tan D.-X., Reiter R.J. Melatonin: An ancient molecule that makes oxygen metabolically tolerable. J. Pineal Res. 2015;59:403–419. doi: 10.1111/jpi.12267. [ DOI ] [ PubMed ] [ Google Scholar ]
- 122. Shi H., Chen K., Wei Y., He C. Fundamental issues of melatonin-mediated stress signaling in plants. Front. Plant Sci. 2016;7:1124. doi: 10.3389/fpls.2016.01124. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 123. Zhang Y.P., Yang S.J., Chen Y.Y. Effects of melatonin on photosynthetic performance and antioxidants in melon during cold and recovery. Biol. Plant. 2017;61:571–578. doi: 10.1007/s10535-017-0717-8. [ DOI ] [ Google Scholar ]
- 124. Wang S.-Y., Shi X.-C., Wang R., Wang H.-L., Liu F., Laborda P. Melatonin in fruit production and postharvest preservation: A review. Food Chem. 2020;320:126642. doi: 10.1016/j.foodchem.2020.126642. [ DOI ] [ PubMed ] [ Google Scholar ]
- 125. Abdelgawad Z. Improving growth and yield of salt-stressed cowpea plants by exogenous application of Ascobin. Life Sci. J. 2014;11:43–51. [ Google Scholar ]
- 126. Allahveran A., Farokhzad A., Asghari M., Sarkhosh A. Foliar application of ascorbic and citric acids enhanced ‘Red Spur’ apple fruit quality, bioactive compounds and antioxidant activity. Physiol. Mol. Biol. Plants. 2018;24:433–440. doi: 10.1007/s12298-018-0514-7. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 127. Rodriguez-Furlán C., Miranda G., Reggiardo M., Hicks G.R., Norambuena L. High throughput selection of novel plant growth regulators: Assessing the translatability of small bioactive molecules from Arabidopsis to crops. Plant Sci. 2016;245:50–60. doi: 10.1016/j.plantsci.2016.01.001. [ DOI ] [ PubMed ] [ Google Scholar ]
- 128. Fozard S., Forde B.G. Novel micro-phenotyping approach to chemical genetic screening for increased plant tolerance to abiotic stress. In: Fauser F., Jonikas M., editors. Plant Chemical Genomics. Volume 1795. Springer; New York, NY, USA: 2018. pp. 9–25. Methods in Molecular Biology. [ DOI ] [ PubMed ] [ Google Scholar ]
- View on publisher site
- PDF (1.1 MB)
- Collections
Similar articles
Cited by other articles, links to ncbi databases.
- Download .nbib .nbib
- Format: AMA APA MLA NLM
Add to Collections
IMAGES
COMMENTS
Plant resistance to abiotic stresses at the molecular level through sensing mechanisms and genetic responses. Abiotic stress can be perceived in different cellular compartments, including the cell wall (CW), plasma membrane (PM), cytoplasm, mitochondria, chloroplasts, peroxisomes, endoplasmic reticulum, and nucleus, leading to the initiation of molecular responses.
Nov 20, 2023 · Plants have undergone evolutionary changes for hundreds of years under the influence of increasing abiotic and biotic stress due to increasing human activities over the past 200 years that has transformed habitats by adding a variety of new harsh stressors including reduced biodiversity and ecosystem pollution (Fig. 1) (Han, 2019; Westwood et al., 2021).
Sep 24, 2021 · Zhu, J. K. Abiotic stress signaling and responses in plants. Cell 167 , 313–324 (2016). This Review proposes the concept of dispersed stress sensing in various cell parts .
Oct 1, 2024 · The collected papers not only show that nanobiotechnology is a powerful tool for improving plant abiotic stress tolerance, but describe the function of transcription factors in regulating such tolerance. These efforts aim to create robust crops with strong abiotic stress tolerance or the ability to withstand multiple combined stresses.
Abiotic stress is defined as the negative impact of non-living factors on the living organisms in a specific environment. The non-living variable... | Explore the latest full-text research PDFs ...
Jun 30, 2023 · Abiotic stress is the adverse effect of any abiotic factor on a plant in a given environment, impacting plants’ growth and development. These stress factors, such as drought, salinity, and extreme temperatures, are often interrelated or in conjunction with each other. Plants have evolved mechanisms to sense these environmental challenges and make adjustments to their growth in order to ...
The number of publications per year related to systems biology and abiotic stress. Key words used in the search of PubMed included: plant, systems biology, and abiotic stress (including stress sub-terms; e.g. drought or water deficit or dehydration). *The number for the year 2011 was estimated by doubling the 6-month value.
Mar 19, 2024 · The Research Topic Abiotic Stress in Plants: Sustainability and Productivity was dedicated to study the effects of stressful conditions of abiotic origin, such as water and thermal stress, heavy metals and soil salinity, on crops and tree species, with focus on molecular and physio metabolic responses or adjustments.
Nov 17, 2011 · The natural environment for plants is composed of a complex set of abiotic stresses and biotic stresses. Plant responses to these stresses are equally complex. Systems biology approaches facilitate a multi-targeted approach by allowing one to identify regulatory hubs in complex networks. Systems biology takes the molecular parts (transcripts, proteins and metabolites) of an organism and ...
Abiotic stress reduces crop yield. Environmental stress factors, such as heat, cold, drought, salinity, and the presence of heavy metals such cadmium, copper, and chromium, elicit stress responses in plants, including an accumulation of reactive oxygen species (ROS) and reduced photosynthetic activity, which ultimately lower plant growth and thus crop yields.